 Magdalena Joanna Winkiel
Magdalena Joanna Winkiel Szymon Chowański
Szymon Chowański Małgorzata Słocińska
Małgorzata Słocińska- Department of Animal Physiology and Developmental Biology, Institute of Experimental Biology, Faculty of Biology, Adam Mickiewicz University, Poznań, Poland
Cancer is still one of the main causes of death worldwide. For this reason, new compounds that have chemotherapeutic potential have been identified. One such group of substances is Solanaceae glycoalkaloids (GAs). They are natural compounds produced by plants widely used in traditional medicine for healing many disorders. Among others, GAs exhibit significant antitumor properties, for example, a strong inhibitory effect on cancer cell growth. This activity can result in the induction of tumor cell apoptosis, which can occur via different molecular pathways. The molecular mechanisms of the action of GAs are the subject of intensive research, as improved understanding could lead to the development of new cancer therapies. The genetic basis for the formation of neoplasms are mutations in protooncogenes, suppressors, and apoptosis-controlling and repair genes; therefore, substances with antineoplastic properties may affect the levels of their expression or the levels of their expression products. Therapeutic compounds can be applied separately or in combination with other drugs to increase the efficiency of cancer therapy; they can act on the cell through various mechanisms at different stages of carcinogenesis, inducing the process of apoptosis, blocking cell proliferation and migration, and inhibiting angiogenesis. This review summarizes the newest studies on the anticancer properties of solanine (SN), chaconine (CH), solasonine (SS), solamargine (SM), tomatine (TT) and their extracts from Solanum plants.
Introduction
Cancer is one of the most common causes of death globally. In 2020, approximately 19 million new cases of cancer and 10 million deaths due to cancer were registered. The number of cancer cases is expected to increase to more than 28 million over the next 20 years (Sung et al., 2021). In men, the most common neoplasms were lung cancer (14.3%) and prostate cancer (14.1%). In women, breast (24.5%) and colorectal cancer (9.4%) are the most frequently diagnosed. In 2020, the highest percentage of cancer deaths in men was caused by lung cancer (21.5%) and liver cancer (10.5%), while in women, breast cancer (15.5%) and lung cancer (13.7%) were the leading causes of death (Sung et al., 2021). These estimations are based on the GLOBOCAN database prepared by the International Agency for Research on Cancer.
Why is cancer so widespread among people worldwide? The cells of our body grow and multiply during life as a result of division processes that run according to the information contained in the genetic material. The cell cycle of healthy cells stops at the right time. New, young cells replace stem cells, which age and undergo apoptosis. All cell divisions are followed by exact replication of genetic material, resulting in two identical DNA molecules. However, it is possible for various errors to occur in this process. These mistakes can appear spontaneously and may be induced by carcinogens or inherited from parents (Cooper, 2000; Sarkar et al., 2013). These errors lead to mutations in the genetic material and, as a consequence, damaged cells may grow and reproduce in an uncontrolled manner. In a healthy organism, there are many mechanisms of tumor suppression. For example, they enable the cell cycle of mutated cells to be stopped. However, if the repair mechanisms do not work properly, a tumor can develop. With age, these systems function less and less efficiently; in addition, DNA damage accumulates in cells during life; therefore, the risk of developing tumors increases over time (Cooper, 2000; Kamal et al., 2022).
The mechanisms of tumor formation are complex, but the direct causes are mutations in genes encoding proteins crucial for the proper course of cell division, such as protooncogenes (stimulating cell proliferation), suppressor genes (inhibiting cell proliferation), genes controlling cell apoptosis, and repair genes. Protooncogenes include genes that encode growth factors and their receptors, messenger proteins, transcription factors, and cell cycle regulators. Mutations may change the structure or quantity of proteins and thus affect the physiological function they perform in a healthy organism (Domagala, 2007; Sarkar et al., 2013; Kamal et al., 2022).
Unlike normal cells, mutant cells divide excessively in an uncontrolled manner and do not fulfill the role of their origin. Benign tumor cells usually respond well to treatment because they are localized in the tissue, e.g., lipoma in fat tissue. However, when tumor cells can travel via the blood or lymphatic system to other parts of the body, we call it a cancer that is a threat to the proper functioning of the body.
Currently, the main method of treating malignant tumors is chemotherapy, a method based on the use of cytostatic drugs that inhibit cell division due to DNA damage or cell cycle inhibition. These drugs are not very specific and damage healthy, rapidly dividing cells such as the bone marrow and hair follicles, causing a number of side effects such as anemia and baldness. Many patients undergo radiation therapy, which results in the production of free radicals that destroy cell DNA. Hormone therapy is used to treat tumors whose growth depends on the stimulating or inhibitory action of certain hormones to produce an antitumor effect, e.g., in breast and thyroid cancer (National Cancer Institute, 2017). Neoplastic lesions can be surgically removed with a margin of healthy tissue. Most modern cancer treatments include immunotherapy and molecularly targeted drugs. In 2018, the Nobel Prize for Physiology or Medicine was awarded to James Allison and Tasuku Honjo for developing cancer immunotherapy based on stimulating the immune system to recognize and destroy cancer cells, e.g., by giving a vaccine. The targeted drugs are monoclonal antibodies or kinase inhibitors, whose action is based on inhibition of the molecular mechanisms of tumor formation and growth (National Cancer Institute, 2017).
Searching for new methods in cancer treatment is critical for global cancer control. It is important to remember that plants through millions years of evolution developed and produce an enormous number of biologically active substances. That is why modern medicine often reaches for achievements of traditional knowledge about medicinal plants built over the years. The first antimalarial drug was obtained from Cinchona spp. Artemisinin, lactone with antimalarial properties, was isolated from Artemisia annua L. The source of morphine was Papaver somniferum L. There are much more examples of medicinal plants usage (Pirintsos et al., 2022). Solanaceae plants are also rich in bioactive metabolites such as alkaloids, glycosides and lignans. Antimicrobial, insecticidal and antiinfectious properties of Solanum plants were used traditionally, for example, pepper (Capsicum annuum L.) was applied to prevent cold and improve digestion, and potato (Solanum tuberosum L.) in case of burns, cough, and spasms (Afroz et al., 2020). Local inhabitants of rural areas around the world have comprehensive traditional knowledge about the use of Solanaceae as medicinal plants for cancer treatment (Khan et al., 2021). However, the path from the traditional ethnopharmacology to discovery of botanical drugs and development a specific method of treatment is not easy to follow, but early descriptions of therapeutic properties of natural plant products can indicate potential application of such compounds in medicine and facilitate developing effective drugs. Moreover, medicinal plants are still often used as an alternative to modern medicine or as primary remedies in developing countries (Vandebroek, 2013; Pirintsos et al., 2022).
Many compounds of plant origin show anticancer properties, such as components of Solanum nigrum L. fruit. They were identified, and numerous cancer-related targets of their action were recognized based on the molecular docking method (Yang et al., 2021). For example, alkaloids, which are alkaline substances with high biological activity that contain a nitrogen atom in their structure, are currently used in cancer therapy. Vinca L. alkaloids (vinblastine and vincristine) and yew (Taxus baccata L.) alkaloids (paclitaxel) act by binding to microtubules that form the mitotic spindle, resulting in inhibition of cell division. Vinblastine and paclitaxel are used in the treatment of breast and lung cancer, and vincristine is used in the treatment of acute lymphoblastic leukemia (Pantziarka et al., 2021). Another compound with anticancer properties is camptothecin, a quinoline alkaloid of Camptotheca acuminata Decne. a topoisomerase I inhibitor. It was withdrawn from clinical trials due to its high toxicity and low water solubility; however, derivatives of this alkaloid are used in the treatment of ovarian and lung cancer (Gurgul and Litynska, 2017).
Glycoalkaloids (GAs), or alkaloids with an attached sugar group, also exhibit toxic properties against cancer cells. One group of plants that is rich in GAs are the nightshades (Solanaceae). They are common in the world and comprise many useful plant species, i.e., potato Solanum tuberosum L., tomato Solanum lycopersicum L., black nightshade S. nigrum L. and sweet pepper Capsicum annuum L. Nightshade plants are a source of many biologically active substances, including alkaloids. For example, atropine present in deadly nightshade (Atropa bella-donna L.) acts as an antagonist of muscarinic receptors, and nicotine occurring in tobacco (Nicotiana tabacum L.) is an agonist of N-acetylcholine nicotinic receptors (Gopalakrishnakone et al., 2017). GAs are secondary metabolites that are sugar derivatives of steroidal alkaloids. GAs have an amphiphilic nature and are composed of a hydrophilic carbohydrate chain attached to the 3-OH position and the aglycone part, which is a 27-carbon skeleton of cholastane with a nitrogen atom in the ring structure. Solanine (SN) and chaconine (CH) are composed of an aglycon called solanidine, the aglycon part of solasonine (SS) and solamargine (SM) is solasodine, and the structure of tomatine (TT) is based on the tomatidine moiety (Milner et al., 2011). The chemical structures of the GAs that are the subject of this study are shown in Figure 1. Most GAs are solids, less frequently liquids, and are well soluble in organic solvents, with high biological activity. These compounds are produced in flowers, leaves, roots, and sprouts. For example, the source of SN is the potato tuber, whereas TT is found in the leaves of tomatoes. GA synthesis in plants occurs during seed germination and is most intense during the flowering period. As a result of exposure to light, potato tubers turn green, and the SN content increases tenfold, as the synthesis of the steroid GAs is related to the biosynthesis of chlorophyll (Friedman, 2006).
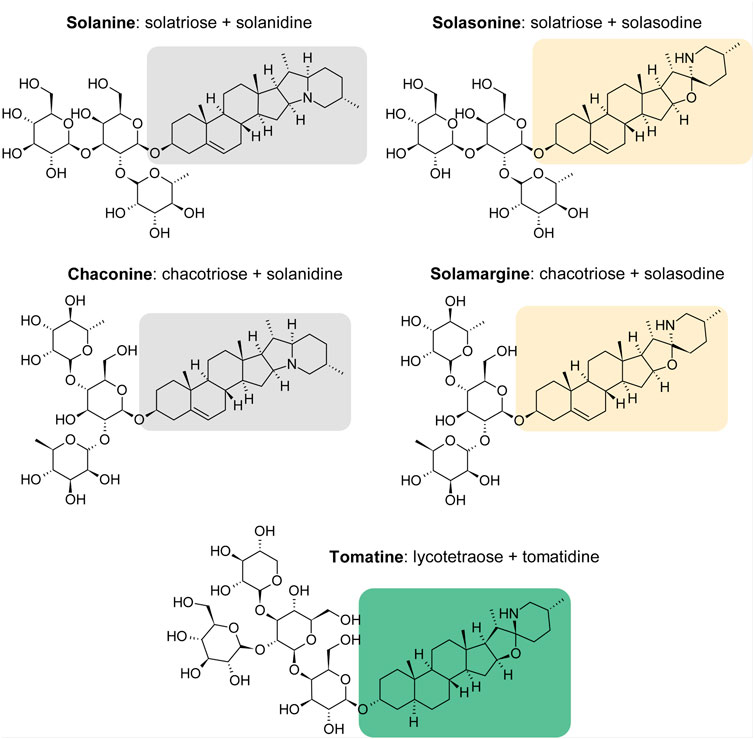
FIGURE 1. Structures of Solanaceae glycoalkaloids.
The molecular mechanisms underlying GA action are the subject of intensive research, as improved understanding could lead to the development of new cancer therapies. The aim of this study is to review the latest research on the anticancer properties of Solanaceae GAs. Studies in English from 2015 to 2022 were reviewed on the Web of Science using the keywords Solanaceae, glycoalkaloids, solanine, chaconine, solasonine, solamargine, tomatine, tumor, cancer and their combinations.
Toxicity of Solanaceae glycoalkaloids
In plants, alkaloids are a natural defense against phytopathogens and herbivores. Pure substances and their plant extracts show dose-dependent toxicity against fungi, insects, and other organisms (Boulogne et al., 2012; Chowanski et al., 2016; Chowanski et al., 2018; Spochacz et al., 2018), and have been used in the production of pharmaceuticals due to their properties (Oksman-Caldentey, 2007). Because ‘only the dose makes a substance a poison’, alkaloids can be therapeutic as well as toxic.
In in silico studies, the toxicity of solanidine, solasodine, SN, SS and SM was calculated and compared with the toxicity of doxorubicin hydrochloride (an anticancer drug) and tetracycline. The analysis predicted that the mentioned GAs do not cause any risk of mutagenic, tumorigenic, or irritant toxicity, although most of them exhibit a mild undesired impact on the reproductive system (Ahmad, 2019). SN influenced oocyte maturation and disrupted embryonic development in a pig model (Lin et al., 2018) and exhibited cytotoxic activity in TM3 and TM4 mouse testis cell lines (Park et al., 2019). SN at a concentration of 20 µM and higher also showed cytotoxicity in human trophoblast cells of the HTR-8/SVneo line. SN increases the expression of autophagic genes, triggers autophagosomal generation, and stops the cell cycle in the S and G2/M phases. By activating apoptosis and autophagy, it inhibits cell viability, migration, invasion, and blood vessel formation (Chen et al., 2021). SN and CH have also been hypothesized to contribute to the formation of congenital abnormalities of the nervous system, consisting of neural tube defect (NTD) formation and disturbances in closure during fetal development; however, there is no evidence of their action in humans (Cuschieri and Calleja-Agius, 2020).
SS and SM show cytotoxic effects in the normal cell lines L-02, BEAS-2B, HK-2, and MCF-10A (Li et al., 2016). Hepatic toxicity and downregulation of cyp450 genes were found in mice treated with SS, and the toxicity depended on the genetic background (Zhong et al., 2018). The IC50 of SS was higher in AGS and HT-29 cells than in healthy cells (Akter et al., 2015). SM from S. nigrum L. fruits exhibited cytotoxic effects in MGC803, HepG2, and SW480 cancer cell lines; however, the LD50 values were higher than those of the positive control KPT330 (Gu et al., 2018). SM at a concentration greater than 14.2 g/ml and SS greater than 28.8 g/ml showed cytotoxicity in V79 cells. However, these GAs do not cause genotoxic effects or aberrations of the chromosome. They cannot reduce the genotoxic effects of camptothecin and etoposide, but SM and SS decreased DNA damage and the prevalence of chromosomal aberrations caused by methyl methanesulfonate, an alkylating compound (Munari et al., 2022). Furthermore, treatment with SM or SM in nanoparticles of yttrium vanadate functionalized with 3-chloropropyltrimethoxysilane at doses of 5 or 10 mg/kg/day administered subcutaneously for 5 days did not show any apparent systemic toxic effects, nephrotoxicity, or genotoxicity in mice. Furthermore, there was less damage to liver DNA caused by melanoma model B16F10 cells in animals treated with 10 mg/kg/day SM (Furtado et al., 2022). SM exhibited only a slight effect on healthy primary bovine aortic endothelial cells (BAECs), rat fibroblasts, and epithelial cells (Al et al., 2016), and the cytotoxic effect of SM and SN on the NIH3T3 and VERO healthy cell lines was smaller than that on the MCF-7, MDA-MB-231, AGS, and HT-29 cancer cell lines (Akter et al., 2015). The extract of the S. nigrum L. fruit exhibited low cytotoxicity (Yang et al., 2022), while there was no toxic effect in mice orally treated with extracts of Solanum capsicoides All. seeds (2000 mg/kg) (Petreanu et al., 2016).
The effects of different concentrations of TT on gene expression and cell monolayer integrity were also studied. Research showed that TT at concentrations lower than 20 μg/ml was safe in the human colon cell line Caco-2. This compound does not affect monolayer integrity or metabolism and does not induce apoptosis or impact the cell cycle; however, it influences cytokine-mediated signaling genes (Arena et al., 2018). The toxicity of the methanolic extract of tomato leaves was evaluated in rats and did not show any hepatotoxic effects even at the highest dose (1,000 mg/kg b.w.) after 28 days of treatment but influenced lipid metabolism and exhibited immunostimulatory effects (Nguenang et al., 2020). Furthermore, the extract of TT from green tomatoes showed a cytotoxic effect on cancer and normal cells. In breaker-stage vegetables, the action of this GA is believed to be masked by antioxidant substances (Del Giudice et al., 2015). For these reasons, special caution should be considered during the study of the pharmaceutical use of GAs (Li et al., 2016).
However, GAs from the Solanaceae family may have a cytotoxic effect on healthy cells as well as cancer cells. Furthermore, their aglycon parts also have anticancer effects (da Silva et al., 2017; Shen et al., 2017; Akhtar et al., 2018; Gu et al., 2018; Fekry et al., 2019; Ahmad, 2019; Hsieh et al., 2020; Fujimaki et al., 2022; Yu et al., 2020). SN, CH, SS, SM, and TT show antitumor activity against various types of neoplasms. Research results and possible mechanisms of GA activity in cancer cells are shown in Figure 2. Although the mode of action of GAs has been intensively studied, it is not yet fully understood. Both GA extracts from Solanaceae plants and compounds used individually have been shown to be effective. The results of recent research on the anticancer properties of Solanaceae GAs are presented below, taking into account the different mechanisms of action of these substances (Table 1).
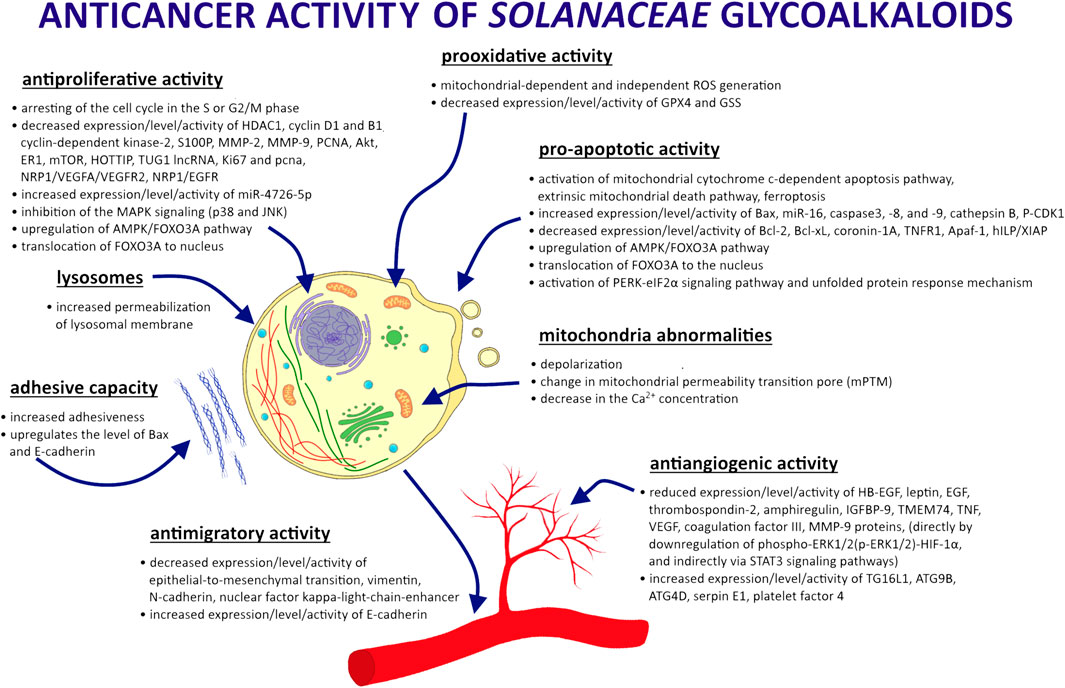
FIGURE 2. Effect of GA action in cancer cells.
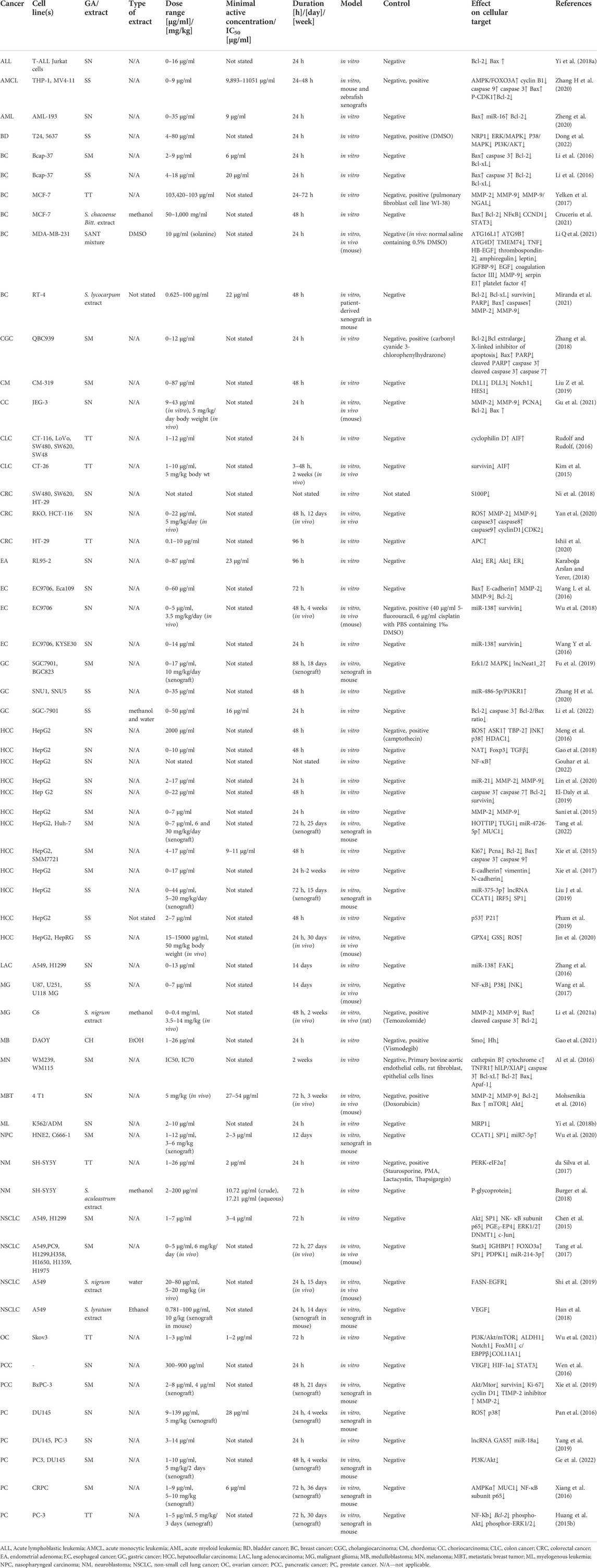
TABLE 1. Effect of GAs on cellular targets in cancer cells.
Antitumor effects of Solanaceae glycoalkaloids
Solanine
Solanine (SN) is one of the two principal S. tuberosum L. GAs, in addition to chaconine (CH), but it also occurs in many other Solanum species. The SN level in potatoes depends on many factors, such as light and temperature. SN is composed of trissacharide solatriose (D-glucose, D-galactose, ʟ-rhamnose) attached to the aglycon part solanidine (Milner et al., 2011). SN exhibits fungicidal (Boulogne et al., 2012), antibacterial (Rajalakshmi and Jayachitra, 2017) and anti-inflammatory activity depending on inhibition of the NF-κB signaling pathway (Shin et al., 2016). SN also exhibits an antitumor effect on different cancer cells.
Most studies of SN anticancer activity were conducted in HepG2 cells of hepatocellular carcinoma (HCC). SN promotes the production of ROS by human HCC HepG2 cells in both mitochondrial-dependent and mitochondrial-independent manners. This compound also upregulates the expression and kinase activity of ASK1 and TBP-2, which are proteins that activate the JNK and p38 signaling pathways. Furthermore, SN decreases the expression level of the proliferation-associated protein HDAC1. Thus, it contributes to cell apoptosis (Meng et al., 2016). Arylamine N-acetyltransferase (NAT) is an enzyme involved in the transformation of arylamines into carcinogens. SN inhibits NAT activity and downregulates its expression in HCC cells (Gao et al., 2018).
The presence of CD4+CD25+ Treg cells in peripheral blood and lymph nodes could be a marker of many types of cancer. The indicator of these cells is a Foxp3 forkhead transcription factor that is involved in the proper functioning of Treg cells. TGFβ is a growth factor of epithelial-derived cells that suppresses the response of the immune system. SN promoted the antitumor immune response by decreasing the proportion of CD4+CD25+ Treg cells and negatively impacting the expression of Foxp3 and TGFβ in mouse HCC cells. The tumor size in the transplanted tumor model in mice was smaller than that in the control (Gao et al., 2020). SN generates oxidative stress in HCC HepG2 cells by upregulation or downregulation of appropriate miRNAs controlling the NF-κB pathway and increasing NF-κB expression (Gouhar et al., 2022, p. 202). SN inhibits the proliferation and migration induced by acetylcholine in HCC Hep G2 cells by attenuating the epithelial-mesenchymal transition. It reduces the activity of matrix metalloproteinases, which play a key role in the development of the neoplastic process and the formation of metastases in tumors of various origins. SN had an inhibitory effect on extrahepatic metastasis by lowering microRNA-21 expression in exosome-treated A549 lung cancer cells (LC) (Lin et al., 2020).
Studies on AML-193 acute myeloid leukemia (AML) cells show that SN causes morphological changes in neoplastic cells and has a dose-dependent pro-apoptotic effect. SN changes the expression of genes that encode the Bax and Bcl-2 proteins, which are markers of apoptosis. SN increases the expression of Bax and miR-16 (noncoding RNA genes that regulate gene expression), resulting in a decrease in Bcl-2 expression, thus inhibiting cell proliferation (Zheng et al., 2020). SN has cytotoxic effects on colorectal cancer (CRC) in vitro and in vivo. It inhibits the growth, migration, and invasion of CRC SW480, SW620, and HT-29 cells and promotes cell cycle arrest as well as apoptosis. Additionally, SN decreases the expression of S100P, which is involved in the regulation of many cellular processes, e.g., cell cycle progression and differentiation. In vivo, this GA inhibits tumor growth (Ni et al., 2018). Akt serine-threonine kinase is the main effector of phosphatidylinositol 3-kinase PI3K, contributing to cell survival by mediating its response to the action of growth factors. Disturbances in the functioning of this transmitter are observed in many disorders, e.g., cancer. SN reduces the expression of genes and the activity of Akt and ER proteins in RL95-2 endometrial adenoma cells (EA) containing the ER estrogen receptor. This may result in inhibition of the PI3K/Akt and ER signaling pathways. The single-administered GA effect was as strong as that of the Akt API-1 inhibitor and the ER-MPP blocker (Karaboğa Arslan and Yerer, 2018). SN inhibits the growth, proliferation, migration, and invasion and promotes apoptosis of human esophageal EC9706 and Eca109 cancer cells (EC) in vitro. It upregulates the levels of Bax and E-cadherin, which regulate the adhesive capacity of cells. However, SN downregulates the expression of MMP-2, MMP-9, and Bcl-2 (Wang L. et al., 2016). SN induces apoptosis in human choriocarcinoma (CC) JEG-3 cells. Moreover, it inhibited proliferation, invasion, and migration in vitro and in vivo in mice. SN decreases MMP-2, MMP-9, proliferative cellular nuclear antigen (PCNA), and Bcl-2 levels, while it increases Bax expression (Gu et al., 2021). SN regulates the expression of cell cycle proteins in prostate cancer (PC) DU145 cells. In addition, it induces apoptosis through the ROS and P38 MAPK kinase signaling pathways. SN also reduces tumor cells in vitro and in vivo (Pan et al., 2016). Under hypoxic conditions, increased expression of crucial regulators of the angiogenesis process was found, such as hypoxia-inducible factor-1 nuclear transcription factor, hypoxia inducible factor-1α (HIF-1α) and phosphorylated signal transducer and activator of transcription 3 (p-STAT3). VEGF is a proangiogenic growth factor that stimulates the formation of new blood vessels, while inhibition of tumor angiogenesis is crucial because it leads to growth inhibition and reduced invasiveness and metastasis formation. SN reduces the expression of VEGF directly by downregulating phospho-ERK1/2(p-ERK1/2)-HIF-1α and indirectly via STAT3 signaling pathways in pancreatic cancer (PCC) cells. Thus, downregulation of VEGF is a combined result of HIF-1α and STAT3 action (Wen et al., 2016). SN exhibited cytotoxic effects in human CRC RKO and HCT-116 cells in vitro and in vivo. It inhibits their proliferation, migration, invasion, and adhesion and induces apoptosis as a result of increased ROS generation and caspase 3, 8, and 9 activation. SN treatment leads to cell cycle arrest in the G0-G1 phase. It decreases the expression levels of cyclin D1 and cyclin-dependent kinase 2. Additionally, SN reduces the expression level and activity of MMP-2 and MMP-9 (Yan et al., 2020). SN showed an even greater cytotoxic effect on human breast cancer (BC) MCF-7 and MDA-MB-231 cells than cycloheximide, which was used as a positive control. Furthermore, SN has an apoptosis-inducing potential and arrests MCF-7 cells in the S phase (Akter et al., 2015). In in silico studies, S. nigrum L. GAs have been identified that may bind to selected cytoskeleton proteins. SN has a binding affinity for coronin-1A, which is a protein that inhibits intrinsic pathway-mediated apoptosis; therefore, inhibiting its function could potentially be a target for anticancer therapy (Ahmad, 2019). SN distributed via biocompatible polymeric carriers called dendrosomes (DNS) to metastatic breast tumor (MBT) 4T1 cells was more efficient in treatment in vitro and in vivo and safer than SN alone. DNS decreased the proliferation of cells and increased the expression of Bax while causing lower expression levels of Bcl-2, MMP-2, MMP-9, mTOR, and Akt (Mohsenikia et al., 2016).
SN also exhibits potential for use in combination with other pharmaceuticals, such as fluorouracil and cisplatin. Fluorouracil is an analog of pyrimidine, which blocks thymidylate synthetase, disrupts DNA synthesis, and blocks the cell cycle in the S phase. Cisplatin is an alkylating agent that attaches alkyl groups to DNA bases, leading to fragmentation of DNA. SN increased the sensitivity of EC9706 and KYSE30 EC cells to these chemotherapeutic agents (Wu et al., 2018). MiR-138 is a microRNA that targets genes associated with proliferation, migration, invasion, and apoptosis. Survivin is a protein involved in regulation of the cell cycle and inhibition of apoptosis. SN induces apoptosis by increasing miR-138 and reducing the expression of the survivin gene. SN also enhanced the action of these cytostatics in EC9706 cells transplanted into mice. MiR-138 inhibitors reverse the effect induced by SN, and overexpression of the gene encoding survivin protects tumor cells from apoptosis (Wu et al., 2018). SN also amplifies the anticancer effect induced by cisplatin in HepG2 HCC cells, such as growth inhibition and induction of apoptosis. The mechanism of action is to stop the cell cycle in the G2/M phase, stimulate DNA fragmentation, and induce cell death by activating caspases 3 and 7. SN also reduces the expression of genes encoding the antiapoptotic proteins Bcl-2 and surviving, and regulates the expression of microRNA-21 (El-Daly et al., 2019). SN also increases the radiosensitivity and chemosensitivity of human lung adenocarcinoma (LAC) A549 and H1299 cancer cell lines by upregulating miR-138 and downregulating focal adhesion kinase (FAK) mRNA and protein expression (Zhang et al., 2016). SN induces apoptosis of acute lymphoblastic leukemia (ALL) human T-ALL Jurkat cells. It increases the level of Bax and reduces Bcl-2 mRNA and protein levels. Furthermore, SN increases the chemosensitivity of these cells to adriamycin (Yi et al., 2018a) and reverses the multidrug resistance of human myelogenous leukemia (ML) K562/ADM cells (Yi et al., 2018b). Treatment with this compound reduces cell proliferation, leads to higher concentrations of adriamycin in cells, and reduces the expression of the MRP1 protein dependent on the JNK pathway (Yi et al., 2018b). SN also increases the radiosensitivity of human PC and EC cells. This compound causes upregulation of the lncRNA GAS5, leading to negative regulation of miR-18a expression by target binding in PC cells (Yang et al., 2019). SN upregulates miR-138 expression in EC cells, leading to decreased expression of survivin genes and increased radiotherapy efficacy (Wang Y. et al., 2016).
Chaconine
Chaconine (CH) was identified in S. tuberosum L. in 1954. The SN and CH ratios in potatoes can differ because they vary, e.g., between cultivars. Chaconine contains solanidine as an aglycone unit and a chacotriose carbohydrate chain (D-galactose and two moieties of ʟ-rhamnose) (Milner et al., 2011). This GA is a bioactive compound with fungicidal activity (Boulogne et al., 2012). Furthermore, CH has been shown to have anti-inflammatory properties in vitro and in vivo and potency for use in the treatment of sepsis (Lee et al., 2015).
There are only a few recent studies on the anticancer effects of CH. Its role in the inhibition of Hh-dependent tumors, such as medulloblastoma (MB), has been determined. This compound reduces Hh pathway activity by binding to the Smo protein. Furthermore, CH exhibits an antiproliferation effect and decreases the growth of MB DAOY cells. The viability of cells treated with CH was even lower than that of cells treated with 22-NHC hydrochloride, a new-generation Smo inhibitor (Gao et al., 2021). CH also decreases the expression of the genes encoding Akt (serine-threonine protein kinase) and ERα (estrogen receptor α) and their activity in the RL95-2 estrogen receptor-positive human endometrial cancer cell line. Thus, it seems that CH may exert suppressive effects on the PI3K/Akt and ERα signaling pathways (Karaboğa Arslan and Yerer, 2018).
Solamargine
Solamargine (SM) can be found in the aubergine Solanum melongena L., but it was also isolated from other Solanaceae species. Solasodine is its hydrophobic unit, and it has the same carbohydrate chain as SN (D-glucose, D-galactose, and ʟ-rhamnose) (Milner et al., 2011). In addition to anticancer activity, SM also exhibits insecticidal and fungicidal effects (Boulogne et al., 2012).
SM is the most active compound of the GAs isolated from the fruit of S. melongena L. It shows cytotoxicity against HCC Huh7 and HepG2 liver cancer cells. The antiproliferative properties of this GA are due to its ability to arrest the cell cycle in the S phase. This compound shows significant pro-apoptotic properties in both of the above cancer cell lines (Fekry et al., 2019). SM significantly reduces HepG2 HCC cell migration and invasion in vitro by decreasing the expression levels of the MMP-2 and MMP-9 genes and reducing the activity of related proteins (Sani et al., 2015). SM also acts through the HOTTIP-TUG1/miR-4726-5p/MUC1 signaling pathway, which inhibits the growth of HCC HepG2 and Huh-7 cells in vitro and in vivo. The compound downregulates the expression levels of HOTTIP and TUG1 lncRNA and upregulates the expression of miR-4726-5p, which inhibits the expression of the MUC1 protein. Furthermore, this GA enhances the antitumor effect of sorafenib, a drug used in HCC treatment (Tang et al., 2022). SM exhibits an antitumor effect in HHC cells, SMM7721 cells, and HepG2 cells. It inhibits proliferation and induces apoptosis and cell cycle arrest in the G2/M phase. SM also reduces the levels of proteins involved in proliferation (Ki67 and pcna) and the antiapoptotic protein Bcl-2. Moreover, it increases the activity of the pro-apoptotic proteins Bax, caspase 3 and caspase 9 (Xie et al., 2015) and suppresses the migration and invasion of HCC HepG2 cells through the inhibition of epithelial-to-mesenchymal transition (EMT), which is necessary for tumor growth. SM upregulates the expression of the EMT-associated epithelial marker E-cadherin and downregulates the expression of the mesenchymal markers vimentin and N-cadherin (Xie et al., 2017).
SM exhibits cytotoxic effects on human BC Bcap-37 cells; it caused apoptosis and depolarization of mitochondria after treatment for 24 h. SM increases the expression levels of Bax and caspase 3 and downregulates the expression levels of Bcl-2 and Bcl-xL. SM induces the mitochondrial cytochrome c-dependent apoptosis pathway (Li et al., 2016). SM showed an even greater cytotoxic effect on human BC MCF-7 and MDA-MB-231 cells than cycloheximide, which was used as a positive control (Akter et al., 2015). SM exhibits cytotoxic activity against breast adenocarcinoma (BAC) MDA-MB-231, LC A549, HCC Hep3B, and PC PC3 cancer cell lines (Tai et al., 2018). Research on mouse melanoma (MN) model B16F10 cells discovered that SM inhibits proliferation and decreases tumor size (Furtado et al., 2022). This glycoalkaloid also suppresses the growth of MN WM239 and WM115 cells. It promotes permeabilization of the lysosomal membrane. SM increases the level of cathepsin B, which contributes to the release of cytochrome c and upregulation of TNFR1, leading to the extrinsic mitochondrial death pathway. In addition, SM downregulates hILP/XIAP, which contributes to caspase 3 cleavage, upregulation of Bcl-xL and Bcl2, and downregulation of Apaf-1 and Bax proteins in cells. Thus, the classic intrinsic apoptosis pathway is probably not involved in the mechanisms of action of this compound (Al et al., 2016). SM suppresses the growth of human non-small cell lung cancer (NSCLC) by influencing the phosphatidylinositol 3-kinase/Akt (PI3-K/Akt) signaling pathway in vitro and in vivo. Prostaglandin E2 (PGE2) belongs to the protein family and is a ligand for G-coupled receptors involved in processes such as growth and metastasis. Inhibition of Akt phosphorylation by GA causes decreased expression of SP1 and NK-κB subunit p65 transcription factors, which contribute to inhibition of the expression of the PGE2 E-prostanoid receptor 4 (EP4) protein (Chen et al., 2015). Subsequently, it was found that decreased EP4 expression results in enhanced ERK1/2 signaling, leading to downregulation of DNA methyltransferase 1 (DNMT1) and c-Jun transcription factor expression; DNMT1 is an enzyme that regulates cell survival, cell cycle arrest, and cell death, and c-Jun is a subunit of activating protein-1 (AP-1) that activates oncogenes (Chen et al., 2017). Moreover, SM inactivates Stat3, causing upregulated expression of the IGHBP1 gene that leads to correlative modulation of FOXO3a (+) and SP1 (-). These interactions caused decreased growth and induce cell cycle arrest in NSCLC cells in vitro and in vivo. Additionally, SM enhances the anticancer effect caused by metformin (Tang et al., 2017). SM inhibited NSCLC cell growth in vitro and in vivo by regulating the interaction between antisense RNA lncRNA HOX transcript (HOTAIR) and miR-214-3p. It results in a downregulation of the expression of the 3-phosphoinositide-dependent protein kinase-1 (PDPK1) gene (Tang et al., 2019).
SM suppresses gastric cancer (GC) progression by decreasing the phosphorylation of extracellular signal-regulated kinase (Erk) 1/2 mitogen-activated protein kinase (MAPK). In this way, the compound induces the expression of the long noncoding RNA nuclear paraspeckle assembly transcript 1 (lncNeat1_2). Furthermore, SM inhibits GC growth in vivo (Fu et al., 2019). SM downregulates the PI3K/Akt pathway and suppresses PC growth in vitro and in vivo. This signaling pathway seems to be responsible for the chemoresistance of this disease. Furthermore, it increases the anticancer effect caused by docetaxel; thus, it may have the potential to be used in therapy together with anticancer drugs (Ge et al., 2022). SM activates AMPKα, reducing the protein expression of MUC1 and NF-κB subunit p65 in vitro in castration-resistant PC CRPC cells and inhibiting tumor growth in vivo. Furthermore, it acts synergistically with metformin (Xiang et al., 2016). SM also shows an antitumor effect in CM-319 human chordoma (CM) cells. It causes cell cycle arrest in the G1 phase, inhibiting proliferation and inducing apoptosis. SM also significantly reduced the expression levels of DLL1, DLL3, Notch1, and HES1, modulating the Notch pathway (Liu J. et al., 2019). SM inhibits the growth of nasopharyngeal carcinoma (NPC) cells in vitro and in vivo by modulating the expression of colon cancer-associated transcript-1 (CCAT1) of lncRNA and miR7-5p. It downregulates CCAT1 and SP1 protein expression and upregulates miR7-5p expression levels (Wu et al., 2020, p. 201). SM induces apoptosis, arrests the cell cycle, inhibits PCC metastasis in vitro, and suppresses PCC growth in vivo. SM acts through inhibition of the Akt/mTOR signaling pathway. The compound downregulates the expression of survivin, Ki-67, and cyclin D1. SM also decreases MMP-2 and increases the expression of metallopeptidase inhibitor-2 of TIMP (Xie et al., 2019). PCC treatment with SM delivered to cells in Fe3O4 magnetic liposomes (MLP) enabled a slow release of the GA and was more effective than SM applied alone. For example, MLPs-SM inhibited Ki-67 expression and caused increased apoptosis compared to SM (Xie et al., 2019). SM induces apoptosis of human cholangiocarcinoma (CGC) QBC939 cells through the mitochondrial pathway and changes the mitochondrial membrane potential in vitro. This compound downregulates the gene expression of Bcl-2, an extralarge and X-linked inhibitor of apoptosis, but upregulates the expression of the Bax gene. Furthermore, SM decreases the level of PARP proteins while increasing the levels of cleaved PARP, caspase 3, cleaved caspase 3 and caspase 7 (Zhang et al., 2018).
Cancer stem cells (CSCs) seem to be the main factors responsible for cancer drug resistance. In silico studies have shown that SM can target CSCs by modifying the sonic hedgehog (SHH) pathway. It also has good binding ability to the Gli protein, which is a transcription factor that mediates transcription related to cancer and CSCs (Mayank, 2016). Furthermore, SM was used as a component in the optimization of new nanoparticles with a bioresponsive off-coating property and an active tumor-targeting ability. These particles have longer circulation, increased accumulation, and penetration efficacy in the cancer area (Li et al., 2021a).
Solasonine
Solasonine (SS) has the same aglycon part as SM (solasodine), and its carbohydrate chain is identical to that of SN (solatriose). It occurs in many Solanum species, such as S. melongena L, S. berthaultii Hawkes, S. platanifolium Sims, and S. ambosinum Ochoa (Milner et al., 2011). SS has insecticidal activity, e.g., against G. mellonella larvae (Spochacz et al., 2021).
Similar to SM, SS can target CSCs by modifying the Hh pathway (Mayank, 2016). The SHH (sonic hedgehog) signaling pathway plays an important role in the growth of neoplastic tumors and in maintaining their resistance to treatment. Stimulation of the SHH pathway increases the expression of genes encoding Patched (PTCH1) and Smoothened (SMO) proteins. PTCH1 binds to the SHH protein, and SMO is responsible for activation of the signaling cascade leading to translocation of GLI transcription factors into the cell nucleus and activation of the transcription of SHH-dependent genes such as HOX, WNT, FGF-4, VEGF, CAPN1, and NRP, causing cell proliferation (Yang et al., 2016). One of the mechanisms of resistance to treatment is the presence of Smo proteins; therefore, their inhibitors are being tested. Research indicates the possibility of using SS in anticancer therapies because it reduces the resistance of cancer cells to the currently used SMO inhibitors. SS significantly inhibits the activity of the SHH pathway as a result of its interaction with the transcription factor Gli. SS inhibits ALP (alkaline phosphatase) in murine C3H10T1/2 mesenchymal cells and reduces the expression of the Gli1 and Ptch1 genes but does not affect the activity of transcription factors induced by TNF-α (tumor necrosis factor α) cytokines and PGE2 prostaglandin, indicating the selective action of SS on cells (Yang et al., 2016).
SS may inhibit the growth of HCC cells through regulatory interactions between miR-375-3p, lncRNA CCAT1, transcription factor SP1, and interferon regulatory factor IRF5, as demonstrated in vivo in a mouse model. This GA increases miR-375-3p expression and decreases levels of lncRNA CCAT1, and consequently leads to SP1-mediated reduction of IRF5 expression, which participates in the regulation of the expression of genes involved in the immune response (Liu J. et al., 2019). Mortalin is a protein that is overexpressed in cancer cells by blocking the action of the transcription factor p53. The arrest of this signaling pathway activates the apoptosis process in HepG2 HCC cells, which has been confirmed in vitro. In silico studies have shown that SS is a potential inhibitor of the p53-mortalin interaction. Furthermore, SS-induced cell death has been shown to occur in cells that do not express the p53 protein, indicating that this process can also be regulated by p53-independent signaling pathways (Pham et al., 2019). Often, the target of anticancer therapies is microtubules (Jordan and Wilson, 2004). In silico studies show the highest binding affinity for coronin-1A of SS among all identified S. nigrum L. GAs, higher than SN (Ahmad, 2019). SS isolated from S. melongena L. fruit shows cytotoxicity against HCC Huh7 and HepG2 liver cancer cells. The antiproliferative properties of this GA are due to its ability to arrest the cell cycle in the S phase. The compound has a significant pro-apoptotic effect on Huh7 cells. (Fekry et al., 2019). SS from S. nigrum L. fruits also exhibited cytotoxic effects in MGC803, HepG2 and SW480 cancer cell lines, however, the LD50 values were higher than that of the positive control KPT330 (Gu et al., 2018). The marker of ferroptosis, which is a form of cell death, is an iron-dependent accumulation of lipid peroxides. In the protection mechanism against this process, glutathione peroxidase 4 (GPX4) is involved, and converts lipid peroxides into nontoxic lipid alcohols. To create a reduced form of GSH, which is the cofactor of GPX4, glutathione synthetase GSS is involved; SS inhibits the expression of GPX4 and GSS, increasing ROS levels and inducing ferroptosis in HepG2 and HepRG cells. The results of research indicate that this GA inhibits proliferation both in vivo and in vitro. SS arrests the cell cycle in the G2/M phase and suppresses the migration and invasion of HCC cells (Jin et al., 2020).
In addition, SS inhibits the proliferation and migration of malignant glioma (MG) human U87 MG, U251 MG and U118 MG cells by reducing the activity of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathway. SS also inhibits the signaling of mitogen-activated protein kinases (MAPKs), namely, p38 and JNK (c-Jun N-terminal kinases). MAPKs are protein kinases that regulate the response to external signals reaching the cell (mitogens) and therefore have an impact on gene expression, division, differentiation, migration, and cell apoptosis (Wang et al., 2017). The latest research shows that SS suppresses neuropilin-1 NRP1 expression in bladder cancer (BD) cells by binding to the b1 domain of the protein on the cell membrane. The NRP1/VEGFA/VEGFR2 and NRP1/EGFR complexes cannot be created, which leads to inhibition of the ERK/MAPK, P38/MAPK, and PI3K/AKT signaling pathways. Furthermore, NRP1 is retained in the cytoplasm, facilitating its degradation. Therefore, this GA probably acts in two ways: intra- and extracellularly (Dong et al., 2022). SS exhibited cytotoxic effects in human BC Bcap-37 cells, and caused apoptosis and depolarization of mitochondria after treatment for 24 h. SS increases the expression levels of Bax and caspase 3 and downregulates the expression levels of Bcl-2 and Bcl-xL. SS induces the mitochondrial cytochrome c-dependent apoptosis pathway (Li et al., 2016). SS showed an even greater cytotoxic effect on BC MCF-7 and MDA-MB-231 cells than cycloheximide, which was used as a positive control (Akter et al., 2015). SS also has anticancer properties against GC; it inhibits its growth and progression, and the mechanism of apoptosis depends on the upregulation of the miR-486-5p/Pi3KR1 pathway (Zhang H. et al., 2020). SS shows cytotoxicity on human GC SGC-7901 cells in a dose-dependent manner. It suppresses proliferation to a degree comparable to that of cisplatin and arrests the cell cycle, mainly in the G2 phase. This compound downregulates the Bcl-2 protein, which controls the mitochondrial permeability transition pore (mPTM), causing a decrease in the Ca2+ concentration within the mitochondria, which leads to an increase in the amount of Ca2+ ions in the cell and a reduction in the mitochondrial membrane potential. Therefore, caspase 3 activation and reduction of the Bcl-2/Bax ratio occur, which contribute to cell cycle arrest in phase M. Therefore, SS is believed to induce the mitochondrial apoptotic pathway because, in addition to Bcl-2, it also decreases the level of Bax expression (Li et al., 2022). SS shows antitumor activity against the acute monocytic leukemia (AMCL) cell lines THP-1 and MV4-11. SS upregulates the AMPK/FOXO3A pathway, inducing FOXO3A translocation to the nucleus. It leads to the induction of cell cycle arrest in the G2/M phase because it inhibits the expression of cyclin B1 and apoptosis as a result of promoting the expression of caspase 9 and caspase 3. SS increases Bax and P-CDK1 expression and downregulates Bcl-2 expression. In addition, it inhibited tumor growth in vivo in mice and zebrafish xenograft models (Zhang Y. et al., 2020).
Sevofluran is an inhalant anesthetic that shows adverse effects on the central nervous system, such as neuronal apoptosis and neuroinflammation. SS has the ability to reduce damage in the mouse hippocampal neuron cell line HT22 caused by the anesthetic by activating the AMPK/FoxO3a pathway. Moreover, AMPK inhibition greatly influences the inhibitory effect of SS on apoptosis, oxidative stress and inflammation. Furthermore, this GA decreased sevoflurane-induced learning and memory disorders in vivo in mice (Zhang and Yan, 2022). Comparison of these two studies shows that the mechanisms of apoptosis in AMCL and neuronal cells are different because the upregulation of the AMPK pathway can lead to apoptosis or, in contrast, to apoptosis inhibition.
Tomatine
Tomatine (TT) is a steroidal glycoalkaloid that occurs in large amounts in the leaves of tomatoes (S. lycopersicum L.). It is composed of an aglycon, a hydrophobic part called tomatidine, a hydrophilic carbohydrate chain consisting of two glucose molecules, one galactose and one xylose molecule (lycotetraose), and a polar amine group. TT is a biologically active substance and, in addition to its anticancer properties, has a wide range of activities, such as insecticidal (Ventrella et al., 2016), fungicidal (Boulogne et al., 2012), antibiotic (Friedman, 2002) and anti-inflammatory (Zhao et al., 2015) effects.
TT at a concentration of 10 μg/ml inhibited CRC HT-29 cell proliferation 24 h after administration but did not induce apoptosis. It increases the expression of the gene encoding the APC protein that is involved in the regulation of the cell cycle (Ishii et al., 2020). Other studies indicate that TT shows pro-apoptotic properties against metastatic melanoma (MSMN) cells. This GA also reduces cell invasion and inhibits angiogenesis, possibly by affecting the ER/p-eIF2/VEGF pathway (Serratì et al., 2020). TT inhibits the growth and apoptosis of human myeloid leukemia (MLL) HL-60 cells in a dose-dependent manner. The cytotoxic effect is based on its ability to bind to cholesterol and, consequently, disrupt the cell membrane and lead to necrosis. TT also inhibited the growth of xenografts of these cancer cells in mice (Huang et al., 2015a). This GA acts through a caspase-independent mechanism (Del Giudice et al., 2015). This was confirmed by another study on the SH-SY5Y neuroblastoma (NM) cell line. TT- and tomatidine-induced cell death has been demonstrated to be independent of caspases and receptor-interacting protein 1 (RIP1) threonine kinase because exposure to the pan Z-VAD-caspase inhibitor fmk and necrostatin-1 (an inhibitor of RIP1 kinase) did not prevent this process. Furthermore, both compounds increase the level of calcium ions in the cell. The increasing number of misfolded proteins in the endoplasmic reticulum activates the UPR (unfolded protein response) stress response mechanism and the activation of receptors: PERK (protein kinase RNA-like endoplasmic reticulum kinase), IRE1 (inositol-requiring-enzyme 1) and ATF6 (activating transcription Factor 6). As a result of the action of TT and tomatidine, the PERK-eIF2α signaling pathway is activated, but the IRE1α pathway is not. Furthermore, TT is an inhibitor of the 20S proteasome, so it influences the concentration of proteins in the cell (da Silva et al., 2017). The increased level of ROS causes overexpression of the gene encoding mitogen-activated protein kinase 6 (MAPK6), which leads to the creation of neoplasms. In silico molecular docking experiments have shown that TT may suppress the expression of MAPK6/ERK3 proteins and protect cells against cancer induced by oxidative stress (Kalimuthu et al., 2021).
TT suppresses proliferation and inhibits autophagy to induce apoptosis in Skov3 cells of ovarian cancer (OC), which was confirmed by measuring the expression of the Beclin-1 gene. Additionally, this GA inhibits the PI3K/Akt/mTOR pathway (Wu et al., 2021). TT induces necroptosis and caspase-independent apoptosis in human colon cancer (CLC) cells, especially those with a hyperactivated PI3K/AKT pathway. This GA changes the expression of genes that encode the RIP3 and RIP1 proteins and causes the accumulation of mitochondrial cyclophilin D. The effects of TT treatment include lysosomal membrane permeabilization and mitochondrial secretion of apoptosis-inducing factor (AIF), which lead to cell death probably through the JNK pathway (Rudolf and Rudolf, 2016). TT induces the death of CLC CT-26 cells through caspase-independent pathways in vitro and in vivo. It promotes the nuclear translocation of AIF and downregulates survivin gene expression. TT generated approximately 50% lysis of cancer cells 24 h after application at a concentration of 3.5 µM (Kim et al., 2015). Matrix metalloproteinases (MMPs) secreted from tumor cells are key factors in neoplasm metastasis. Research shows that TT increased apoptosis and suppressed the activation of MMP-2, MMP-9 and MMP-9/NGAL in the human BC cell line MCF-7 treated with this compound (Yelken et al., 2017).
TT may also have the potential to be used in combined therapy. Porphyrin complexes of platinum are used in photodynamic therapy, where singlet oxygen is formed under the influence of light, which results in the death of tumor cells. Because of its chemical properties, TT creates self-assembled nanostructured supramolecules. Therefore, it could be used in combined therapy with Pt complexes as it improves their incorporation into cells and increases the efficiency of photodynamic therapy (Fujitsuka et al., 2021). In addition, TT and curcumin used in combination inhibited growth more effectively and promoted apoptosis in PC-3 cells in vitro. These substances synergistically suppress the activity of NF-κB and decrease the expression of Bcl-2. The TT-curcumin effect was also observed as decreased levels of phospho-Akt and phospho-ERK1/2 in cancer cells. This combined therapy also slowed cancer cell growth in vivo (xenografts in mice) (Huang et al., 2015b).
Crude extracts of Solanaceae plants
GA extracts of Solanaceae plants have also been studied due to their anticancer properties. The ripe fruit extract of S. nigrum L. shows potent antitumor properties in vitro (LC A549 cells) and in vivo in mice with Lewis tumors in a dose-dependent manner. Cancer cells show increased expression of genes encoding fatty acid synthase (FASN), which correlates with resistance to anticancer drugs targeting the epidermal growth factor receptor (EGFR). Both S. nigrum L. extract and the newly identified GA of this plant, solaoiacid, inhibit the FASN-EGFR signaling pathway and affect the migration, autophagy, apoptosis, and immune response of non-small cell lung cancer (NSCLC) cells. Furthermore, solaoiacid acts with an IC50 = 2.3 μmol/L, which is significantly lower than that of SM (Shi et al., 2019). The extract of S. nigrum L. had antitumor effects on rat C6 MG in vitro and in vivo. It inhibits the viability, proliferation, migration, and invasion of cancer cells. Moreover, the extract induced annexin V + PI + late-stage apoptosis and decreased the protein levels of MMP-2 and MMP-9. It also upregulates Bax and cleaved caspase 3 and downregulates the expression of Bcl-2. In rat brains, the extract of S. nigrum L. inhibited the proliferation, growth, and penetration of cancer tissue (Li et al., 2021a). The methanolic extracts of the dried root, stem, and leaves of S. nigrum L. diminished proliferation and promoted apoptosis in the human cancer cell lines MDA and A549 and, particularly, HepG2. Healthy Vero cells were significantly vulnerable to the extract. Moreover, molecular docking analysis shows S. nigrum L. GA binding affinity to vimentin, which is thought to be a marker of epithelial-mesenchymal transition and is crucial for cancer metastasis (Ahmad et al., 2017). Vimentin belongs to the group of intermediate filament (IF) proteins, which are one of the three systems of the cell cytoskeleton, in addition to actin and myosin filaments and microtubules. IFs are highly flexible and have an α-helical conformation. Vimentin IFs are responsible for the structural properties of cells as well as functional processes. For example, these proteins control signaling pathways, inflammatory responses and even metabolism (Herrmann and Aebi, 2016; Ridge et al., 2022).
Solanum aculeastrum Dunal fruit extract, containing SM and SN, had a cytotoxic effect and inhibited the activity of P-glycoprotein in both neoplastic and healthy cells. SH-SY5Y NM cells (NM) were more susceptible to the effects of both the crude extract and its water fraction; cytotoxic effects were observed, and inhibition of P-glycoprotein activity was caused by the action of SM (Burger et al., 2018).
Wild potato leaf extract Solanum chacoense Bitt., containing solasodine, SM, SN and CH, exhibited selective cytotoxic properties for MCF7 breast cancer BC cells compared to healthy HUVECs (human umbilical vein endothelial cells). Both the leaf extract and the tuber extract of this plant increased the expression of the pro-apoptotic Bax gene and decreased the expression of the anti-apoptotic Bcl-2 gene and genes promoting the proliferation of NFκB, CCND1, and STAT3 cells. For this reason, GAs or other substances such as phenolic acids may be responsible for the properties of these extracts (Cruceriu et al., 2021).
Actinic keratosis is a pathological change that predisposes patients to squamous cell carcinoma of the skin. SR-T100 is a newly patented drug extracted from Solanum incanum L. It contains SM as an active ingredient and is used in the form of a gel. In the third phase of clinical trials, the effectiveness and safety of its use in the treatment of actinic keratosis was demonstrated (Yang et al., 2018). This drug increases the expression of tumor necrosis factor and activates the mitochondrial apoptosis pathway. The IC50 for SR-T100 is higher in OC cells than in healthy cells. The drug decreased the expression of aldehyde dehydrogenase 1 (ALDH1), Notch1, FoxM1, c/EBPPβ and COL11A1 genes. Furthermore, SR-T100 suppressed the growth of A2780CP70 cells in mouse xenografts and, in combination with cisplatin, increased the effectiveness of ovarian cancer treatment (Wu et al., 2015). SR-T100 has been shown to induce apoptosis, DNA degradation, and cell cycle arrest in the G0/G1 phase in murine B16 MN cells. Additionally, in in vivo studies in mice, intravascular injection of the drug reduced tumor size, and intraperitoneal injection inhibited LC growth and the number of metastases (Yu et al., 2017).
The extract of S. melongena L. peel applied twice a day for 12 weeks reduced the diameter of skin lesions in arsenic-induced Bowen disease (Sarah and Misbahuddin, 2018).
SN is one of the four components of the SANT mixture, which contains active ingredients of traditional Chinese botanical drugs that exhibit anticancer effects. The potential of SANT in treating heparanase (HPSE)-related triple-negative breast cancer (TNBC) was identified in vitro and in vivo. HPSE is a glucuronidase responsible for the resistance of tumor cells to chemotherapy. It promotes proliferation, invasion, metastasis, and angiogenesis of the tumor. SANT upregulates the expression of genes encoding ATG16L1, ATG9B, and ATG4D while downregulating TMEM74 and TNF gene expression. Moreover, it decreases the levels of HB-EGF, thrombospondin-2, amphiregulin, leptin, IGFBP-9, EGF, coagulation factor III and MMP-9 proteins but increases serpin E1 and platelet Factor 4 levels. The mixture of botanical drugs also reduces tumor growth and angiogenesis in vivo (Li et al., 2021b).
To enhance the anticancer effect of GAs, high temperature may be used. Research shows that the effect of high temperature on S. tuberosum L. juice increases the antioxidant potential and cytotoxic properties toward Caco-2 and HT-29 intestinal cancer (IT) cells. At the same time, the cytotoxicity of this extract for nonmutated cells is the same or lower than the cytotoxicity of fresh juice. Thermal deproteinization does not affect the content of GAs in the juice, while spray-drying decreases in its content. Low temperatures do not affect the GA content and do not increase the cytotoxic properties against neoplastic cells (Kowalczewski et al., 2019).
Furthermore, the extract of SM and SS from S. lycocarpum A.St.-Hil. has been shown to cause chemosensitization of RT-4 BC cells to cisplatin chemotherapy, indicating a synergistic effect of both substances. PDX cells (patient-derived xenografts) were characterized by much higher resistance to cisplatin than RT-4 cells. PDX xenografts are human models composed of cancer cells introduced into the body of immunodeficient mice and are used to test anticancer drugs. The extract in combination with cisplatin induced apoptosis and inhibited the growth and migration of RT-4 cells. The mechanism of apoptosis induced by cisplatin and GA extract is a decrease in the expression of genes encoding anti-apoptotic proteins Bcl-2, Bcl-xL, survivin, and PARP and increase in the expression of genes encoding the pro-apoptotic protein Bax and activate caspases. Proteins of the PARP family (poly (ADP-ribose) polymerase) are responsible for the repair of damaged DNA; therefore, excessive expression of the genes encoding them may make cancer cells resistant to treatment. Furthermore, a decrease in the expression of MMP-2 and MMP-9 metalloproteinases was observed, indicating the effect of this combination therapy on cell invasion and migration (Miranda et al., 2021).
Because of the poor solubility of GAs in water and low bioavailability, delivery of GA to cancer cells is an issue. An-te-xiao capsules containing alkaloids obtained from whole dried S. lyratum Thunb. plants had an inhibitory effect on NSCLC cells from the A549, NCI-H460 and NCI-H520 lines in vitro and in vivo. These capsules significantly extended the life of mice with Lewis lung cancer cells (xenografts). An-te-xiao influences tumor growth, microvessel density, cell cycle regulatory proteins, and apoptosis and causes histopathological changes in neoplastic cells. In vitro studies showed that the capsules inhibited migration, invasion, and vessel formation in Td-ECs (tumor-derived endothelial cells) both in the presence and absence of VEGF. Additionally, they decrease VEGF secretion by A549 lung adenocarcinoma cells and phosphorylation of this growth factor receptor (Han et al., 2018). GA nanocapsules with a diameter of 125 ± 6 nm were also created using polylactic acid, and the cytotoxic effect of the S. lycocarpum extract was observed in RT-4 BD cells. Apoptosis and blocking of the cell cycle in the S phase were demonstrated; therefore, the use of nanocapsules may overcome the poor solubility of GAs in water (Miranda et al., 2019). The use of natural lipid-based nanoparticles (myristyl myristate, illipe butter) also decreased the viability of bladder cancer cells by 5.4 times (Carvalho et al., 2019). Additionally, nanoparticles with a particle size of 177 nm functionalized with folate were developed and used against BC MDA-MB-231 and BD RT4 cancer cells. The IC50 for these nanoparticles was lower than that of the free extract, indicating that they can penetrate cancer cells. Furthermore, nanoparticles have greater cellular uptake in cancer cells than healthy cells (Miranda et al., 2020). These results suggest that the use of GA-loaded nanoparticles is promising in cancer treatment and could increase the therapeutic efficacy of applied glycoalkaloids.
Conclusion and future perspectives
Solanaceae GAs seem to be a promising agent for the development of alternative methods of cancer treatment. This study summarizes the latest in silico, in vitro, and in vivo studies on the anticancer properties of these compounds, with a focus on their mechanisms of action. These plant secondary metabolites have the potential to become new pharmaceuticals for the treatment of tumors because they act on different cancer cell lines, even at low concentrations. GAs are involved in many cell signaling pathways; therefore, they can influence cancer cells through diverse mechanisms, such as interactions with microtubules or changes in Bax and Bcl-2 expression. These compounds may be applied separately or in combination with other drugs or therapies, improving their effectiveness. GA extracts from plants also have promising cytotoxic effects on tumor cells due to the synergistic effects of their components. Key factors that influence the anticancer activity of GAs include the location, type, and number of carbohydrate molecules. There is a strong relationship between structure and function; therefore, modification of GA structures may also allow for the discovery of new potential anticancer drugs (Akter et al., 2015; Ahmad, 2019). However, some studies indicate that cytotoxic effects due to these compounds also occur in healthy cells. Therefore, in the search for new directions for the use of GAs, it is important to consider the safety of GA drugs. Taken together, although the effects of GAs on cells have been intensively studied, the precise mechanisms of their action in organisms remain to be elucidated. More research needs to be conducted to explore new, more selective, and less toxic cancer therapies and improve the effectiveness of applied treatment methods because cancer remains one of the leading causes of death worldwide.
Author contributions
All authors contributed to the preparation of this review. MW contributed to the writing and prepared the figures. SC contributed to the writing, prepared the figures and worked to finalize the manuscript. MS reviewed the manuscript and approved the final version. The manuscript was approved by all coauthors.
Acknowledgments
MW thanks SC and MS for their guidance, valuable suggestions, and support to complete this project.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Afroz, M., Akter, S., Ahmed, A., Rouf, R., Shilpi, J. A., Tiralongo, E., et al. (2020). Ethnobotany and antimicrobial peptides from plants of the Solanaceae family: An update and future prospects. Front. Pharmacol. 11, 565–615. doi:10.3389/fphar.2020.00565
Ahmad, R., Gupta, A., Fatima, A., Husain, I., and Srivastava, A. N. (2017). The evaluation of biological activity of methanolic extracts of Solanum nigrum and molecular docking analysis of selected phytoconstituents against vimentin. J. Intercult. Ethnopharmacol. 6, 391–400. doi:10.5455/jice.20170701065821
Ahmad, R. (2019). Steroidal glycoalkaloids from Solanum nigrum target cytoskeletal proteins: An in silico analysis. PeerJ 7, e6012. doi:10.7717/peerj.6012
Akhtar, S., Khan, M. K. A., and Arif, J. M. (2018). Evaluation and elucidation studies of natural aglycones for anticancer potential using apoptosis-related markers: An in silico study. Interdiscip. Sci. 10, 297–310. doi:10.1007/s12539-016-0191-6
Akter, R., Uddin, S. J., Tiralongo, J., Grice, I. D., and Tiralongo, E. (2015). A new cytotoxic steroidal glycoalkaloid from the methanol extract of Blumea lacera leaves. J. Pharm. Pharm. Sci. 18, 616–633. doi:10.18433/J3161Q
Al, S., Sana, S., Eltayeb, E. A., and Coomber, B. L. (2016). Solamargine triggers cellular necrosis selectively in different types of human melanoma cancer cells through extrinsic lysosomal mitochondrial death pathway. Cancer Cell Int. 16, 11. doi:10.1186/s12935-016-0287-4
Arena, M. P., Govers, C., Lotti, C., Ricciardi, L., Wichers, H. J., and Mes, J. J. (2018). The effect of tomatine on gene expression and cell monolayer integrity in Caco-2. Molecules 23, E644. doi:10.3390/molecules23030644
Boulogne, I., Petit, P., Ozier-Lafontaine, H., Desfontaines, L., and Loranger-Merciris, G. (2012). Insecticidal and antifungal chemicals produced by plants: A review. Environ. Chem. Lett. 10 (4), 325–347. doi:10.1007/s10311-012-0359-1
Burger, T., Mokoka, T., Fouché, G., Steenkamp, P., Steenkamp, V., and Cordier, W. (2018). Solamargine, a bioactive steroidal alkaloid isolated from Solanum aculeastrum induces non-selective cytotoxicity and P-glycoprotein inhibition. BMC Complement. Altern. Med. 18, 137. doi:10.1186/s12906-018-2208-7
Carvalho, I. P. S., Miranda, M. A., Silva, L. B., Chrysostomo-Massaro, T. N., Paschoal, J. A. R., Bastos, J. K., et al. (2019). In vitro anticancer activity and physicochemical properties of Solanum lycocarpum alkaloidic extract loaded in natural lipid-based nanoparticles. Colloid Interface Sci. Commun. 28, 5–14. doi:10.1016/j.colcom.2018.11.001
Chen, Y., Tang, Q., Wu, J., Zheng, F., Yang, L., and Hann, S. S. (2015). Inactivation of PI3-K/Akt and reduction of SP1 and P65 expression increase the effect of solamargine on suppressing EP4 expression in human lung cancer cells. J. Exp. Clin. Cancer Res. 34, 154. doi:10.1186/s13046-015-0272-0
Chen, Y., Tang, Q., Xiao, Q., Yang, L., and Hann, S. (2017). Targeting EP4 downstream C-Jun through ERK1/2-mediated reduction of DNMT1 reveals novel mechanism of solamargine-inhibited growth of lung cancer cells. J. Cell. Mol. Med. 21 (2), 222–233. doi:10.1111/jcmm.12958
Chen, Z., Li, C., Yuan, A., Gu, T., Zhang, F., Fan, X., et al. (2021). α-Solanine causes cellular dysfunction of human trophoblast cells via apoptosis and autophagy. Toxins 13, 67. doi:10.3390/toxins13010067
Chowanski, S., Adamski, Z., Marciniak, P., Rosinski, G., Buyukguzel, E., Buyukguzel, K., et al. (2016). A review of bioinsecticidal activity of Solanaceae alkaloids. Toxins 8, 60. doi:10.3390/toxins8030060
Chowanski, S., Chudzinska, E., Lelario, F., Ventrella, E., Marciniak, P., Miadowicz-Kobielska, M., et al. (2018). Insecticidal properties of Solanum nigrum and Armoracia rusticana extracts on reproduction and development of Drosophila melanogaster. Ecotoxicol. Environ. Saf. 162, 454–463. doi:10.1016/j.ecoenv.2018.07.030
Cooper, G. M. (2000). The development and causes of cancer. 2nd edition. (Sunderland, MA: Sinauer Associates).
Cruceriu, D., Diaconeasa, Z., Socaci, S., Socaciu, C., Balacescu, O., and Rakosy-Tican, E. (2021). Extracts of the wild potato species Solanum chacoense on breast cancer cells: Biochemical characterization, in vitro selective cytotoxicity and molecular effects. Nutr. Cancer 73, 630–641. doi:10.1080/01635581.2020.1761407
Cuschieri, A., and Calleja-Agius, J. (2020). Wnt/PCP signalling cascade disruption by JNK inhibition as a potential mechanism underlying the teratogenic effects of potato glycoalkaloids. Mol. Biol. Rep. 47 (11), 9235–9238. doi:10.1007/s11033-020-05921-6
da Silva, D. C., Andrade, P. B., Valentão, P., and Pereira, D. M. (2017). Neurotoxicity of the steroidal alkaloids tomatine and tomatidine is RIP1 kinase- and caspase-independent and involves the EIF2α branch of the endoplasmic reticulum. J. Steroid Biochem. Mol. Biol. 171, 178–186. doi:10.1016/j.jsbmb.2017.03.009
Del Giudice, R., Raiola, A., Tenore, G. C., Frusciante, L., Barone, A., Monti, D. M., et al. (2015). Antioxidant bioactive compounds in tomato fruits at different ripening stages and their effects on normal and cancer cells. J. Funct. Foods 18, 83–94. doi:10.1016/j.jff.2015.06.060
Domagala, W. (2007). Molekularne podstawy karcynogenezy i ścieżki sygnałowe niektórych nowotworów ośrodkowego układu nerwowego. Pol. Przegląd Neurol. 3, 127–141.
Dong, Y., Hao, L., Shi, Z., Fang, K., Yu, H., Zang, G., et al. (2022). Solasonine induces apoptosis and inhibits proliferation of bladder cancer cells by suppressing NRP1 expression. J. Oncol. 2022, 7261486–7261515. doi:10.1155/2022/7261486
El-Daly, S. M., Gouhar, S. A., Gamal-Eldeen, A. M., Abdel Hamid, F. F., Ashour, M. N., and Hassan, N. S. (2019). Synergistic effect of α-solanine and cisplatin induces apoptosis and enhances cell cycle arrest in human hepatocellular carcinoma cells. Anticancer. Agents Med. Chem. 19, 2197–2210. doi:10.2174/1871520619666190930123520
Fekry, M. I., Ezzat, S. M., Salama, M. M., Alshehri, O. Y., and Al-Abd, A. M. (2019). Bioactive glycoalkaloides isolated from Solanum melongena fruit peels with potential anticancer properties against hepatocellular carcinoma cells. Sci. Rep. 9, 1746. doi:10.1038/s41598-018-36089-6
Friedman, M. (2002). Tomato glycoalkaloids: Role in the plant and in the diet. J. Agric. Food Chem. 50, 5751–5780. doi:10.1021/jf020560c
Friedman, M. (2006). Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 54, 8655–8681. doi:10.1021/jf061471t
Fu, R., Wang, X., Hu, Y., Du, H., Dong, B., Ao, S., et al. (2019). Solamargine inhibits gastric cancer progression by regulating the expression of LncNEAT1_2 via the MAPK signaling pathway. Int. J. Oncol. 54 (5), 1545–1554. doi:10.3892/ijo.2019.4744
Fujimaki, J., Sayama, N., Shiotani, S., Suzuki, T., Nonaka, M., Uezono, Y., et al. (2022). The steroidal alkaloid tomatidine and tomatidine-rich tomato leaf extract suppress the human gastric cancer-derived 85As2 cells in vitro and in vivo via modulation of interferon-stimulated genes. Nutrients 14, 1023. doi:10.3390/nu14051023
Fujitsuka, M., Iohara, D., Oumura, S., Matsushima, M., Sakuragi, M., Anraku, M., et al. (2021). Supramolecular assembly of hybrid Pt(II) porphyrin/tomatine analogues with different nanostructures and cytotoxic activities. ACS Omega 6, 13284–13292. doi:10.1021/acsomega.1c01239
Furtado, R. A., Ozelin, S. D., Ferreira, N. H., Miura, B. A., Junior, S. A., Magalhaes, G. M., et al. (2022). Antitumor activity of solamargine in mouse melanoma model: Relevance to clinical safety. J. Toxicol. Environ. Health. A 85 (4), 131–142. doi:10.1080/15287394.2021.1984348
Gao, S., Tan, H., Lang, L., Bai, J., and Ji, Y. (2018). The effect of alpha-solanine on the activity, gene expression, and kinetics of arylamine N-acetyltransferase in HepG2 cells. Oncol. Rep. 39 (5), 2427–2435. doi:10.3892/or.2018.6315
Gao, J., Ying, Y., Wang, J., and Cui, Y. (2020). Solanine inhibits immune escape mediated by hepatoma Treg cells via the tgfβ/smad signaling pathway. Biomed. Res. Int. 9749631, 9749631. doi:10.1155/2020/9749631
Gao, L. J., Zhang, M. Z., Li, X. Y., Huang, W. K., Xu, S. F., and Ye, Y. P. (2021). Steroidal alkaloids isolated from Veratrum grandiflorum Loes. as novel Smoothened inhibitors with anti-proliferation effects on DAOY medulloblastoma cells. Bioorg. Med. Chem. 39, 116166. doi:10.1016/j.bmc.2021.116166
Ge, J., Wang, P., Ma, H., and Zhang, J. (2022). Solamargine inhibits prostate cancer cell growth and enhances the therapeutic efficacy of docetaxel via Akt signaling. J. Oncol. 9055954, 9055954. doi:10.1155/2022/9055954
Gopalakrishnakone, P., Carlini, C. R., and Ligabue-Braun, R. (2017). Plant toxins. Netherlands: Springer.
Gouhar, S. A., Abo-Elfadl, M. T., Gamal-Eldeen, A. M., and El-Daly, S. M. (2022). Involvement of miRNAs in response to oxidative stress induced by the steroidal glycoalkaloid alpha-solanine in hepatocellular carcinoma cells. Environ. Toxicol. 37 (2), 212–223. doi:10.1002/tox.23390
Gu, X.-Y., Shen, X.-F., Wang, L., Wu, Z.-W., Li, F., Chen, B., et al. (2018). Bioactive steroidal alkaloids from the fruits of Solanum nigrum. Phytochemistry 147, 125–131. doi:10.1016/j.phytochem.2017.12.020
Gu, T., Yuan, W., Li, C., Chen, Z., Wen, Y., Zheng, Q., et al. (2021). Alpha-solanine inhibits proliferation, invasion, and migration, and induces apoptosis in human choriocarcinoma JEG-3 cells in vitro and in vivo. Toxins 13, 210. doi:10.3390/toxins13030210
Gurgul, A., and Litynska, A. (2017). Substancje pochodzenia roślinnego w terapii nowotworów. Postępy Fitoter. 18. doi:10.25121/PF.2017.18.3.203
Han, L., Wang, J. N., Cao, X-Q., Sun, C. X., and Du, X. (2018). An-te-xiao capsule inhibits tumor growth in non-small cell lung cancer by targeting angiogenesis. Biomed. Pharmacother. 108, 941–951. doi:10.1016/j.biopha.2018.09.124
Herrmann, H., and Aebi, U. (2016). Intermediate filaments: Structure and assembly. Cold Spring Harb. Perspect. Biol. 8, a018242. doi:10.1101/cshperspect.a018242
Hsieh, M.-H., Yang, J.-S., Lin, R.-C., Hsieh, Y.-H., Yang, S.-F., Chang, H.-R., et al. (2020). Tomatidine represses invasion and migration of human osteosarcoma U2OS and HOS cells by suppression of Presenilin 1 and c-Raf-MEK-ERK pathway. Molecules 25, 326. doi:10.3390/molecules25020326
Huang, H., Chen, S., Doren, J. V., Li, D., Farichon, C., He, Y., et al. (2015a). α-Tomatine inhibits growth and induces apoptosis in HL-60 human myeloid leukemia cells. Mol. Med. Rep. 11 (6), 4573–4578. doi:10.3892/mmr.2015.3238
Huang, H., Chen, X., Li, D., He, Y., Li, Y., Du, Z., et al. (2015b). Combination of alpha-tomatine and curcumin inhibits growth and induces apoptosis in human prostate cancer cells. Plos One 10, e0144293. doi:10.1371/journal.pone.0144293
Ishii, P. L., Oliveira, R. J., Mauro, M. de O., Zorgetto-Pinheiro, V. A., Sartori, D., Lepri, S. R., et al. (2020). The α-tomatine exhibits antiproliferative activity, rupture of cell membranes and induces the expression of Apc gene in the human colorectal adenocarcinoma cell line (Ht-29). Braz. Arch. Biol. Technol. 63. doi:10.1590/1678-4324-2020190395
Jin, M., Shi, C., Li, T., Wu, Y., Hu, C., and Huang, G. (2020). Solasonine promotes ferroptosis of hepatoma carcinoma cells via glutathione peroxidase 4-induced destruction of the glutathione redox system. Biomed. Pharmacother. 129, 110282. doi:10.1016/j.biopha.2020.110282
Jordan, M. A., and Wilson, L. (2004). Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 4 (4), 253–265. doi:10.1038/nrc1317
Kalimuthu, A. K., Panneerselvam, T., Pavadai, P., Pandian, S. R. K., Sundar, K., Murugesan, S., et al. (2021). Pharmacoinformatics-based investigation of bioactive compounds of rasam (South Indian recipe) against human cancer. Sci. Rep. 11, 21488. doi:10.1038/s41598-021-01008-9
Kamal, N., Ilowefah, M. A., Hilles, A. R., Anua, N. A., Awin, T., Alshwyeh, H. A., et al. (2022). Genesis and mechanism of some cancer types and an overview on the role of diet and nutrition in cancer prevention. Molecules 27, 1794. doi:10.3390/molecules27061794
Karaboğa Arslan, A. K., and Yerer, M. B. (2018). α-Chaconine and α-solanine inhibit RL95-2 endometrium cancer cell proliferation by reducing expression of Akt (Ser473) and ERα (Ser167). Nutrients 10, E672. doi:10.3390/nu10060672
Khan, A., Ali, S., Murad, W., Hayat, K., Siraj, S., Jawad, M., et al. (2021). Phytochemical and pharmacological uses of medicinal plants to treat cancer: A case study from khyber pakhtunkhwa, north Pakistan. J. Ethnopharmacol. 281, 114437. doi:10.1016/j.jep.2021.114437
Kim, S. P., Nam, S. H., and Friedman, M. (2015). The tomato glycoalkaloid alpha-tomatine induces caspase-independent cell death in mouse colon cancer CT-26 cells and transplanted tumors in mice. J. Agric. Food Chem. 63, 1142–1150. doi:10.1021/jf5040288
Kowalczewski, P. Ł., Olejnik, A., Białas, W., Kubiak, P., Siger, A., Nowicki, M., et al. (2019). Effect of thermal processing on antioxidant activity and cytotoxicity of waste potato juice. Open Life Sci. 14, 150–157. doi:10.1515/biol-2019-0017
Lee, K.-G., Lee, S.-G., Lee, H.-H., Lee, H. J., Shin, J.-S., Kim, N.-J., et al. (2015). Alpha-chaconine isolated from a Solanum tuberosum L. cv Jayoung suppresses lipopolysaccharide-induced pro-inflammatory mediators via AP-1 inactivation in RAW 264.7 macrophages and protects mice from endotoxin shock. Chem. Biol. Interact. 235, 85–94. doi:10.1016/j.cbi.2015.04.015
Li, N., Cao, L., Wang, Y., Tao, X., Ding, G., Wang, Z., et al. (2016). Induction of solasonine on apoptosis of human breast cancer Bcap-37 cells through mitochondria-mediated pathway. Chin. Herb. Med. 8, 164–172. doi:10.1016/S1674-6384(16)60026-6
Li, J., Tang, W., Yang, Y., Shen, Q., Yu, Y., Wang, X., et al. (2021a). A programmed cell-mimicking nanoparticle driven by potato alkaloid for targeted cancer chemoimmunotherapy. Adv. Healthc. Mat. 10, 2100311. doi:10.1002/adhm.202100311
Li, J., Li, S.-Y., Shen, M.-X., Qiu, R.-Z., Fan, H.-W., and Li, Y.-B. (2021b). Anti-tumor effects of Solanum nigrum L. extraction on C6 high-grade glioma. J. Ethnopharmacol. 274, 114034. doi:10.1016/j.jep.2021.114034
Li, Q., Zhang, G. L., Hao, C. X., Ma, Y-F., Sun, X., Zhang, Y., et al. (2021). SANT, a novel Chinese herbal monomer combination, decreasing tumor growth and angiogenesis via modulating autophagy in heparanase overexpressed triple-negative breast cancer. J. Ethnopharmacol. 266, 113430. doi:10.1016/j.jep.2020.113430
Li, T.-C., Chen, N.-J., Chen, Y.-Y., He, B.-J., and Zhou, Z.-F. (2022). Solasonine induces apoptosis of the SGC-7901 human gastric cancer cell line in vitro via the mitochondria-mediated pathway. J. Cell. Mol. Med. 26, 3387–3395. doi:10.1111/jcmm.17343
Lin, T., Oqani, R. K., Lee, J. E., Kang, J. W., Kim, S. Y., Cho, E. S., et al. (2018). α-Solanine impairs oocyte maturation and quality by inducing autophagy and apoptosis and changing histone modifications in a pig model. Reprod. Toxicol. 75, 96–109. doi:10.1016/j.reprotox.2017.12.005
Lin, L.-T., Choong, C.-Y., and Tai, C.-J. (2020). Solanine attenuated hepatocarcinoma migration and invasion induced by acetylcholine. Integr. Cancer Ther. 19, 1534735420909895. doi:10.1177/1534735420909895
Liu, J., Wang, Z., Xu, C., Qi, Y., and Zhang, Q. (2019). Solamargine inhibits proliferation and promotes apoptosis of CM-319 human chordoma cells through suppression of Notch pathway. Transl. Cancer Res. 8 (2), 509–519. doi:10.21037/tcr.2019.03.07
Liu, Z., Ma, C., Tang, X., Tang, Q., Lou, L., Yu, Y., et al. (2019). The reciprocal interaction between LncRNA CCAT1 and miR-375-3p contribute to the downregulation of IRF5 gene expression by solasonine in HepG2 human hepatocellular carcinoma cells. Front. Oncol. 9, 1081. doi:10.3389/fonc.2019.01081
Mayank, J. V. (2016). Molecular docking study of natural alkaloids as multi-targeted hedgehog pathway inhibitors in cancer stem cell therapy. Comput. Biol. Chem. 62, 145–154. doi:10.1016/j.compbiolchem.2015.08.001
Meng, X. Q., Zhang, W., Zhang, F., Yin, S. Y., Xie, H. Y., Zhou, L., et al. (2016). Solanine-induced reactive oxygen species inhibit the growth of human hepatocellular carcinoma HepG2 cells. Oncol. Lett. 11, 2145–2151. doi:10.3892/ol.2016.4167
Milner, S. E., Brunton, N. P., Jones, P. W., O’Brien, N. M., Collins, S. G., and Maguire, A. R. (2011). Bioactivities of glycoalkaloids and their aglycones from Solanum species. J. Agric. Food Chem. 59, 3454–3484. doi:10.1021/jf200439q
Miranda, M. A., Marcato, P. D., Carvalho, I. P. S., Letícia, B. S., Ribeiro, D. L., Amaral, R., et al. (2019). Assessing the cytotoxic potential of glycoalkaloidic extract in nanoparticles against bladder cancer cells. J. Pharm. Pharmacol. 7110, 1520–1531. doi:10.1111/jphp.13145
Miranda, M. A., Silva, L. B., Carvalho, I. P. S., Amaral, R., de Paula, M. H., Swiech, K., et al. (2020). Targeted uptake of folic acid-functionalized polymeric nanoparticles loading glycoalkaloidic extract in vitro and in vivo assays. Colloids Surf. B Biointerfaces 192, 111106. doi:10.1016/j.colsurfb.2020.111106
Miranda, M. A., Marcato, P. D., Mondal, A., Chowdhury, N., Gebeyehu, A., Surapaneni, S. K., et al. (2021). Cytotoxic and chemosensitizing effects of glycoalkaloidic extract on 2D and 3D models using RT4 and patient derived xenografts bladder cancer cells. Mat. Sci. Eng. C Mat. Biol. Appl. 119, 111460. doi:10.1016/j.msec.2020.111460
Mohsenikia, M., Farhangi, B., Alizadeh, A. M., Khodayari, H., Khodayari, S., Khori, V., et al. (2016). Therapeutic effects of dendrosomal solanine on a metastatic breast tumor. Life Sci. 148, 260–267. doi:10.1016/j.lfs.2016.02.008
Munari, C. C., Ferreira, N. H., Nicolella, H. D., de Oliveira, P. F., Miranda, M. A., Bastos, J. K., et al. (2022). Effects of glycoalkaloids from Solanum lycocarpum on genomic instability. Rev. Bras. Farmacogn. 32, 273–279. doi:10.1007/s43450-022-00249-7
National Cancer Institute (2017). Cancer treatment. Available at: https://www.cancer.gov/about-cancer/treatment (Accessed May 15, 2022).
Nguenang, G. S., Ntyam, A. S. M., and Kuete, V. (2020). Acute and subacute toxicity profiles of the methanol extract of Lycopersicon esculentum L. leaves (tomato), a botanical with promising in vitro anticancer potential. Evid. Based. Complement. Altern. Med. 8935897, 8935897. doi:10.1155/2020/8935897
Ni, X., Chen, J., Lu, F., Yuan, Z., Xu, X., Hu, Z., et al. (2018). Anti-cancer effect of α-solanine by down-regulating S100P expression in colorectal cancer cells. Recent Pat. anticancer. Drug Discov. 13, 240–247. doi:10.2174/1574892813666180329163945
Oksman-Caldentey, K. M. (2007). Tropane and nicotine alkaloid biosynthesis-novel approaches towards biotechnological production of plant-derived pharmaceuticals. Curr. Pharm. Biotechnol. 8 (4), 203–210. doi:10.2174/138920107781387401
Pan, B., Zhong, W., Deng, Z., Lai, C., Chu, J., Jiao, G., et al. (2016). Inhibition of prostate cancer growth by solanine requires the suppression of cell cycle proteins and the activation of ROS/p38 signaling pathway. Cancer Med. 5 (11), 3214–3222. doi:10.1002/cam4.916
Pantziarka, P., Capistrano I, R., De Potter, A., Vandeborne, L., and Bouche, G. (2021). An open access database of licensed cancer drugs. Front. Pharmacol. 12, 627574. doi:10.3389/fphar.2021.627574
Park, S., Park, M. Y., Song, G., and Lim, W. (2019). Alpha-solanine inhibits cell proliferation via mitochondrial dysfunction and inhibin synthesis in mouse testis in vitro and in vivo. Chemosphere 235, 271–279. doi:10.1016/j.chemosphere.2019.06.172
Petreanu, M., Alves Guimaraes, A. A., Broering, M. F., Ferreira, E. K., Machado, I. D., Tasca Gois, A. L., et al. (2016). Antiproliferative and toxicological properties of methanolic extract obtained from Solanum capsicoides All. seeds and carpesterol. Naunyn. Schmiedeb. Arch. Pharmacol. 389, 1123–1131. doi:10.1007/s00210-016-1275-x
Pham, M. Q., Tran, T. H. V., Pham, Q. L., and Gairin, J. E. (2019). In silico analysis of the binding properties of solasonine to mortalin and p53, and in vitro pharmacological studies of its apoptotic and cytotoxic effects on human HepG2 and Hep3b hepatocellular carcinoma cells. Fundam. Clin. Pharmacol. 33, 385–396. doi:10.1111/fcp.12447
Pirintsos, S., Panagiotopoulos, A., Bariotakis, M., Daskalakis, V., Lionis, C., Sourvinos, G., et al. (2022). From traditional ethnopharmacology to modern natural drug discovery: A methodology discussion and specific examples. Molecules 27, 4060. doi:10.3390/molecules27134060
Rajalakshmi, A., and Jayachitra, A. (2017). Antioxidant, antibacterial effects of solanine isolated from Solanum nigrum and its cytotoxic activity on the HEP-2 and AGS cell lines. Int. J. Pharm. Sci. Res. 8 (7), 2932–2939. doi:10.13040/IJPSR.0975-8232.8(7).2932-39
Ridge, K. M., Eriksson, J. E., Pekny, M., and Goldman, R. D. (2022). Roles of vimentin in health and disease. Genes Dev. 36 (7–8), 391–407. doi:10.1101/gad.349358.122
Rudolf, K., and Rudolf, E. (2016). Antiproliferative effects of alpha-tomatine are associated with different cell death modalities in human colon cancer cells. J. Funct. Foods 27, 491–502. doi:10.1016/j.jff.2016.10.005
Sani, I. K., Marashi, S. H., and Kalalinia, F. (2015). Solamargine inhibits migration and invasion of human hepatocellular carcinoma cells through down-regulation of matrix metalloproteinases 2 and 9 expression and activity. Toxicol. Vitro 29 (5), 893–900. doi:10.1016/j.tiv.2015.03.012
Sarah, Q. S., and Misbahuddin, M. (2018). Effect of Solanum melongena peel extract in the treatment of arsenic-induced Bowen’s disease. Bangladesh J. Pharmacol. 13, 309–315. doi:10.3329/bjp.v13i4.38273
Sarkar, S., Horn, G., Moulton, K., Oza, A., Byler, S., Kokolus, S., et al. (2013). Cancer development, progression, and therapy: An epigenetic overview. Int. J. Mol. Sci. 14, 21087–21113. doi:10.3390/ijms141021087
Serratì, S., Porcelli, L., Guida, S., Ferretta, A., Iacobazzi, R. M., Cocco, T., et al. (2020). Tomatine displays antitumor potential in in vitro models of metastatic melanoma. Int. J. Mol. Sci. 21, E5243. doi:10.3390/ijms21155243
Shen, K. H., Hung, J. H., Chang, C. W., Weng, Y. T., Wu, M. J., and Chen, P. S. (2017). Solasodine inhibits invasion of human lung cancer cell through downregulation of MiR-21 and MMPs expression. Chem. Biol. Interact. 268, 129–135. doi:10.1016/j.cbi.2017.03.005
Shi, F., Wang, C., Wang, L., Song, X., Yang, H., Fu, Q., et al. (2019). Preparative isolation and purification of steroidal glycoalkaloid from the ripe berries of Solanum nigrum L. by preparative HPLC-MS and UHPLC-TOF-MS/MS and its anti-non-small cell lung tumors effects in vitro and in vivo. J. Sep. Sci. 42, 2471–2481. doi:10.1002/jssc.201801165
Shin, J.-S., Lee, K.-G., Lee, H.-H., Lee, H. J., An, H.-J., Nam, J.-H., et al. (2016). Solanine isolated from Solanum tuberosum L. cv Jayoung abrogates LPS-induced inflammatory responses via NF-B inactivation in RAW 264.7 macrophages and endotoxin-induced shock model in mice. J. Cell. Biochem. 117, 2327–2339. doi:10.1002/jcb.25530
Spochacz, M., Chowański, S., Szymczak, M., Lelario, F., Bufo, S. A., and Adamski, Z. (2018). Sublethal effects of Solanum nigrum fruit extract and its pure glycoalkaloids on the physiology of Tenebrio molitor (mealworm). Toxins (Basel) 10, E504. doi:10.3390/toxins10120504
Spochacz, M., Chowanski, S., Szymczak-Cendlak, M., Marciniak, P., Lelario, F., Salvia, R., et al. (2021). Solanum nigrum extract and solasonine affected hemolymph metabolites and ultrastructure of the fat body and the midgut in Galleria mellonella. Toxins 13, 617. doi:10.3390/toxins13090617
Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., et al. (2021). Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca. Cancer J. Clin. 71, 209–249. doi:10.3322/caac.21660
Tai, B. H., Van Doan, V., Yen, P. H., Nhiem, N. X., Cuc, N. T., Trang, D. T., et al. (2018). Two new steroidal alkaloid saponins from the whole plants of Solanum nigrum. Nat. Product. Commun. 13, 1934578X1801301. doi:10.1177/1934578X1801301111
Tang, Q., Zheng, F., Wu, J., QianXiaoLi, L., and Hann, S. S. (2017). Combination of solamargine and metformin strengthens igfbp1 gene expression through inactivation of Stat3 and reciprocal interaction between FOXO3a and SP1. Cell. Physiol. biochem. 43, 2310–2326. doi:10.1159/000484383
Tang, Q., Zheng, F., Liu, Z., Wu, J. J., Chai, X., He, C., et al. (2019). Novel reciprocal interaction of LncRNA HOTAIR and MiR-214-3p contribute to the solamargine-inhibited PDPK1 gene expression in human lung cancer. J. Cell. Mol. Med. 23 (11), 7749–7761. doi:10.1111/jcmm.14649
Tang, Q., Li, X., Chen, Y., Long, S., Yu, Y., Sheng, H., et al. (2022). Solamargine inhibits the growth of hepatocellular carcinoma and enhances the anticancer effect of Sorafenib by regulating HOTTIP-TUG1/MiR-4726-5p/MUC1 pathway. Mol. Carcinog. 61 (4), 417–432. doi:10.1002/mc.23389
Vandebroek, I. (2013). Intercultural health and ethnobotany: How to improve healthcare for underserved and minority communities? J. Ethnopharmacol. 148, 746–754. doi:10.1016/j.jep.2013.05.039
Ventrella, E., Adamski, Z., Chudzinska, E., Miadowicz-Kobielska, M., Marciniak, P., Buyukguzel, E., et al. (2016). Solanum tuberosum and Lycopersicon esculentum leaf extracts and single metabolites affect development and reproduction of Drosophila melanogaster. Plos One 11, e0155958. doi:10.1371/journal.pone.0155958
Wang, L., Sun, Q.-Q., Zhang, S.-J., Du, Y.-W., Wang, Y.-Y., Zang, W.-Q., et al. (2016). Inhibitory effect of solanine on esophageal carcinoma in vitro. Exp. Ther. Med. 12, 1525–1530. doi:10.3892/etm.2016.3500
Wang, Y., Wu, J., Guo, W., Sun, Q., Chen, X., Zang, W., et al. (2016). α-Solanine modulates the radiosensitivity of esophageal cancer cells by inducing MicroRNA 138 expression. Cell. Physiol. biochem. 39, 996–1010. doi:10.1159/000447807
Wang, X., Zou, S., Lan, Y. L., Xing, J. S., Lan, X. Q., and Zhang, B. (2017). Solasonine inhibits glioma growth through anti-inflammatory pathways. Am. J. Transl. Res. 9 (9), 3977–3989.
Wen, Z., Huang, C., Xu, Y., Xiao, Y., Tang, L., Dai, J., et al. (2016). α-Solanine inhibits vascular endothelial growth factor expression by down-regulating the ERK1/2-HIF-1α and STAT3 signaling pathways. Eur. J. Pharmacol. 771, 93–98. doi:10.1016/j.ejphar.2015.12.020
Wu, Y.-H., Chiu, W.-T., Young, M.-J., Chang, T.-H., Huang, Y.-F., and Chou, C.-Y. (2015). Solanum incanum extract downregulates aldehyde dehydrogenase 1-mediated stemness and inhibits tumor formation in ovarian cancer cells. J. Cancer 6, 1011–1019. doi:10.7150/jca.12738
Wu, J., Wang, L., Du, X., Sun, Q., Wang, Y., Li, M., et al. (2018). α-Solanine enhances the chemosensitivity of esophageal cancer cells by inducing microRNA-138 expression. Oncol. Rep. 39 (3), 1163–1172. doi:10.3892/or.2018.6187
Wu, J., Tang, X., Ma, C., Shi, Y., Wu, W., and Hann, S. S. (2020). The regulation and interaction of colon cancer-associated transcript-1 and mir7-5p contribute to the inhibition of sp1 expression by solamargine in human nasopharyngeal carcinoma cells. Phytother. Res. 34 (1), 201–213. doi:10.1002/ptr.6555
Wu, H., Li, W., Wang, T., Rong, Y., He, Z., Huang, S., et al. (2021). α-Tomatine, a novel early-stage autophagy inhibitor, inhibits autophagy to enhance apoptosis via Beclin-1 in Skov3 cells. Fitoterapia 152, 104911. doi:10.1016/j.fitote.2021.104911
Xiang, S. S., Zhang, Q., Tang, Q., Zheng, F., Wu, J., Yang, L., et al. (2016). Activation of AMPK alpha mediates additive effects of solamargine and metformin on suppressing MUC1 expression in castration-resistant prostate cancer cells. Sci. Rep. 6, 36721. doi:10.1038/srep36721
Xie, X., Zhu, H., Yang, H., Huang, W., Wu, Y., Wang, Y., et al. (2015). Solamargine triggers hepatoma cell death through apoptosis. Oncol. Lett. 10 (1), 168–174. doi:10.3892/ol.2015.3194
Xie, X., Zhu, H., Zhang, J., Wang, M., Zhu, L., Guo, Z., et al. (2017). Solamargine inhibits the migration and invasion of HepG2 cells by blocking epithelial-to-mesenchymal transition. Oncol. Lett. 14 (1), 447–452. doi:10.3892/ol.2017.6147
Xie, X., Zhang, X., Chen, J., Tang, X., Wan, M., Zhang, L., et al. (2019). Fe3O4-solamargine induces apoptosis and inhibits metastasis of pancreatic cancer cells. Int. J. Oncol. 54 (3), 905–915. doi:10.3892/ijo.2018.4637
Yan, X., Li, M., Chen, L., Peng, X., Que, Z. J., An, H. M., et al. (2020). α-Solanine inhibits growth and metastatic potential of human colorectal cancer cells. Oncol. Rep. 43 (5), 1387–1396. doi:10.3892/or.2020.7519
Yang, J., Huang, W., and Tan, W. (2016). Solasonine, a natural glycoalkaloid compound, inhibits Gli-mediated transcriptional activity. Molecules 21, 1364. doi:10.3390/molecules21101364
Yang, C. C., Wong, T. W., Lee, C. H., Hong, C. H., Chang, C. H., Lai, F. J., et al. (2018). Efficacy and safety of topical SR-t100 gel in treating actinic keratosis in taiwan: A phase III randomized double-blind vehicle-controlled parallel trial. J. Dermatol. Sci. 90 (3), 295–302. doi:10.1016/j.jdermsci.2018.02.015
Yang, J., Hao, T., Sun, J., Wei, P., and Zhang, H. (2019). Long noncoding RNA GAS5 modulates alpha-solanine-induced radiosensitivity by negatively regulating MiR-18a in human prostate cancer cells. Biomed. Pharmacother. 112, 108656. doi:10.1016/j.biopha.2019.108656
Yang, L., Gao, S., Su, Z., Qin, X., and Li, Z. (2021). Identification of the constituents and the cancer-related targets of the fruit of Solanum nigrum based on molecular docking and network pharmacology. J. Pharm. Biomed. Anal. 200, 114067. doi:10.1016/j.jpba.2021.114067
Yang, Y., Liu, L., Wu, T., Wang, W., Wang, J., and Lin, L. (2022). Two new steroidal alkaloids from the mature fruits of Solanum nigrum. Phytochem. Lett. 48, 81–86. doi:10.1016/j.phytol.2022.02.002
Yelken, B. O., Balcı, T., Süslüer, S. Y., Kayabaşı, C., Avcı, C. B., Kırmızıbayrak, P. B., et al. (2017). The effect of tomatine on metastasis related matrix metalloproteinase (MMP) activities in breast cancer cell model. Gene 627, 408–411. doi:10.1016/j.gene.2017.06.054
Yi, Y., Jia, X. H., Wang, J. Y., Chen, J. R., Wang, H., and Li, Y. J. (2018a). Solanine induced apoptosis and increased chemosensitivity to adriamycin in T-cell acute lymphoblastic leukemia cells. Oncol. Lett. 15 (5), 7383–7388. doi:10.3892/ol.2018.8229
Yi, Y., Jia, X. H., Zhu, C., Wang, J. Y., Chen, J. R., Wang, H., et al. (2018b). Solanine reverses multidrug resistance in human myelogenous leukemia K562/ADM cells by downregulating MRP1 expression. Oncol. Lett. 15, 10070–10076. doi:10.3892/ol.2018.8563
Yu, S., Sheu, H. M., and Lee, C. H. (2017). Solanum Incanum extract (SR-T100) induces melanoma cell apoptosis and inhibits established lung metastasis. Oncotarget 861, 103509–103517. doi:10.18632/oncotarget.21508
Yu, T., Wu, Q., You, X., Zhou, H., Xu, S., He, W., et al. (2020). Tomatidine alleviates osteoporosis by downregulation of p53. Med. Sci. Monit. 26, e923996. doi:10.12659/MSM.923996
Zhang, H., and Yan, L. (2022). Solasonine relieves sevoflurane-induced neurotoxicity via activating the AMP-activated protein kinase/FoxO3a pathway. Hum. Exp. Toxicol. 41, 9603271211069984. doi:10.1177/09603271211069984
Zhang, F., Yang, R., Zhang, G., Cheng, R., Bai, Y., Zhao, H., et al. (2016). Anticancer function of alpha-solanine in lung adenocarcinoma cells by inducing microRNA-138 expression. Tumour Biol. 37 (5), 6437–6446. doi:10.1007/s13277-015-4528-2
Zhang, X., Yan, Z., Xu, T., An, Z., Chen, W., Wang, X., et al. (2018). Solamargine derived from Solanum nigrum induces apoptosis of human cholangiocarcinoma QBC939 cells. Oncol. Lett. 15, 6329–6335. doi:10.3892/ol.2018.8171
Zhang, H., Tian, F., Jiang, P., Qian, S., Dai, X., Ma, B., et al. (2020). Solasonine suppresses the proliferation of acute monocytic leukemia through the activation of the AMPK/FOXO3A axis. Front. Oncol. 10, 585297. doi:10.3389/fonc.2020.585297
Zhang, Y., Han, G., Cao, Y., Zhang, Y., Zhang, X., and Gong, H. (2020). Solasonine inhibits gastric cancer proliferation and enhances chemosensitivity through MicroRNA-486-5p. Am. J. Transl. Res. 12 (7), 3522–3530.
Zhao, B., Zhou, B., Bao, L., Yang, Y., and Guo, K. (2015). Alpha-tomatine exhibits anti-inflammatory activity in lipopolysaccharide-activated macrophages. Inflammation 38, 1769–1776. doi:10.1007/s10753-015-0154-9
Zheng, Y., Li, L., Gao, Q., Niu, B., and Wang, H. (2020). Solanine inhibits proliferation and promotes apoptosis of the human leukemia cells by targeting the MiR-16/Bcl-2 axis. J. Buon 25 (3), 1614–1618.
Keywords: Solanaceae, glycoalkaloids, Solanum plants, natural compounds, antitumor potential, cancer, molecular mechanisms, apoptosis
Citation: Winkiel MJ, Chowański S and Słocińska M (2022) Anticancer activity of glycoalkaloids from Solanum plants: A review. Front. Pharmacol. 13:979451. doi: 10.3389/fphar.2022.979451
Received: 27 June 2022; Accepted: 08 November 2022;
Published: 07 December 2022.
Edited by:
Michael Heinrich, University College London, United KingdomReviewed by:
Karuppaiya Palaniyandi, Bharathidasan University, IndiaXiao Ding, Kunming Institute of Botany (CAS), China
Copyright © 2022 Winkiel, Chowański and Słocińska. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Magdalena Joanna Winkiel, magwin@amu.edu.pl