
Attenuation of Cigarette-Smoke-Induced Oxidative Stress, Senescence, and Inflammation by Berberine-Loaded Liquid Crystalline Nanoparticles: In Vitro Study in 16HBE and RAW264.7 Cells

 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Formulation and Physiochemical Characterisation of BBR-LCNs
2.2. Cell Culture and Reagents
2.3. Preparation of Cigarette-Smoke Extract (CSE)
2.4. MTT Assay (Cell-Viability Assay)
2.5. Total Cellular Reactive Oxygen Species Assay
2.5.1. Fluorescence Intensity Quantification
2.5.2. Fluorescence Imaging
2.6. Senescence Assay
2.6.1. X-Gal Staining
2.6.2. Immunocytochemistry of p21
2.7. Human Cytokines Protein Array
2.8. Nitric Oxide (NO) Assay
2.9. Real Time-qPCR (Oxidative Stress, Senescence, and Inflammation Gene)
2.10. Statistical Analysis
3. Results
3.1. Preparation and Physicochemical Characterisation of BBR-LCNs Formulation
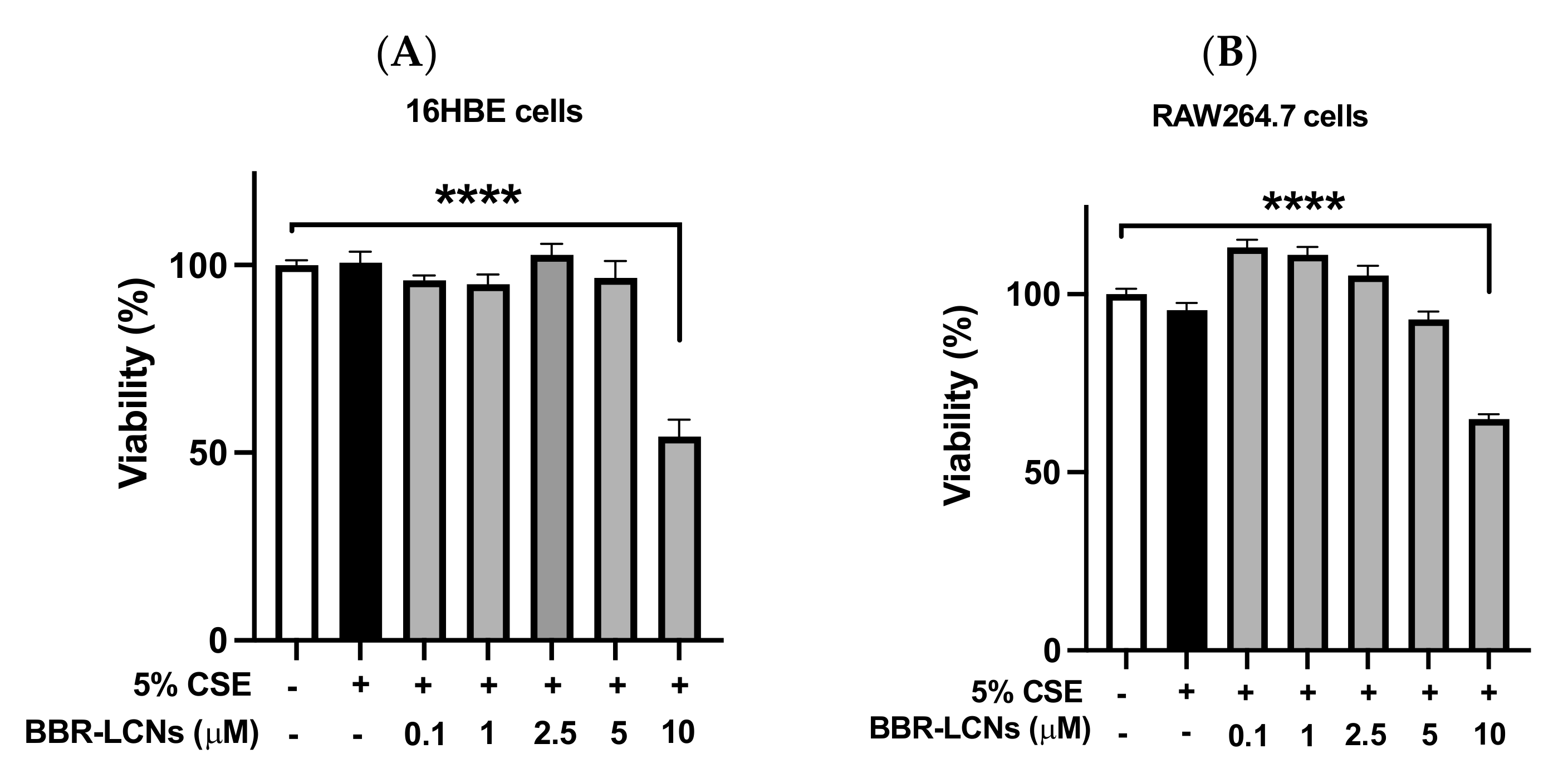
3.2. Viability of BBR-LCNs Treated 16HBE and RAW264.7 Cells
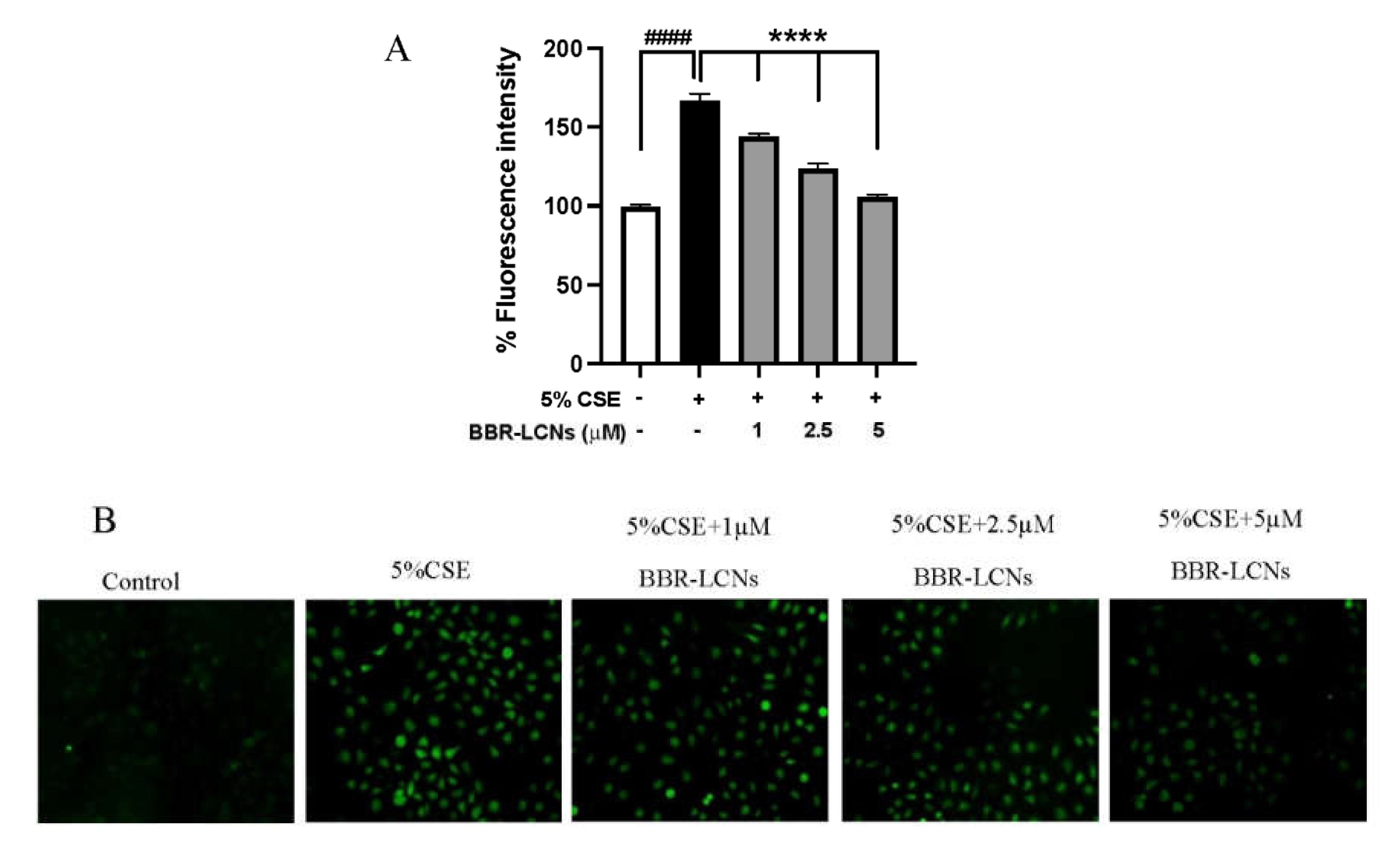
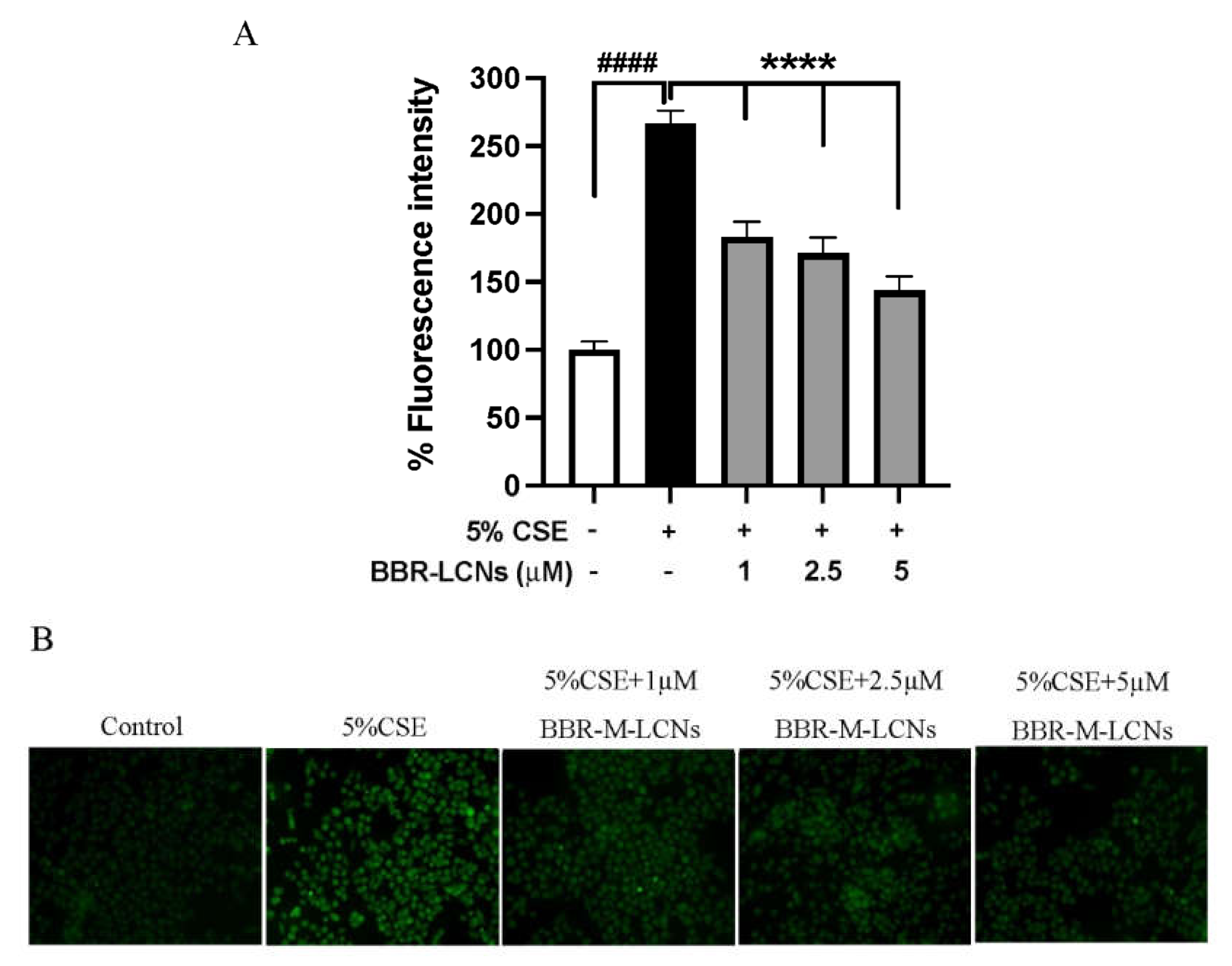
3.3. Inhibition of CSE Induced ROS Generation in 16HBE Cells by BBR-LCNs
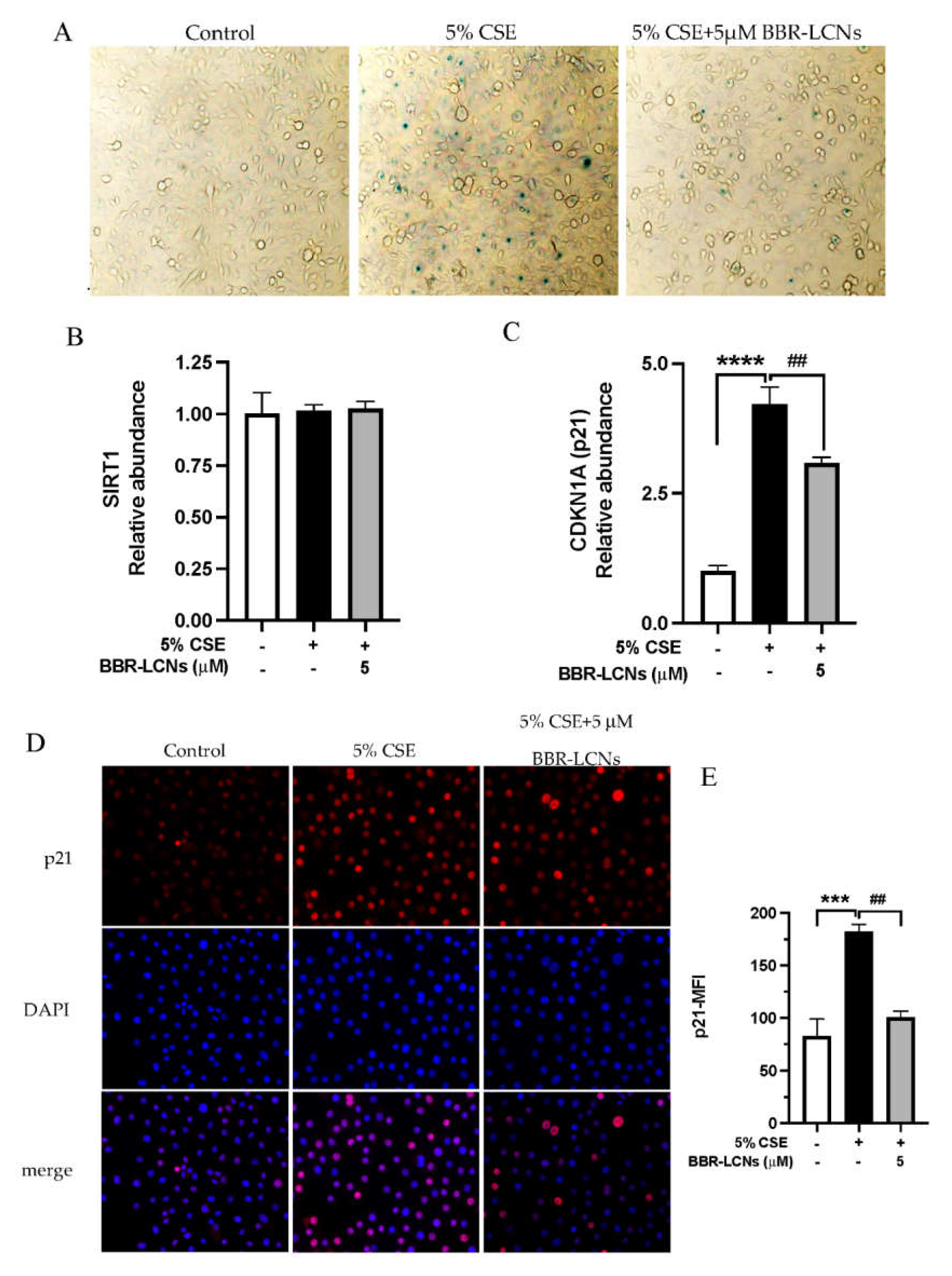
3.4. Inhibition of CSE Induced Senescence of 16HBE Cells by BBR-LCNs
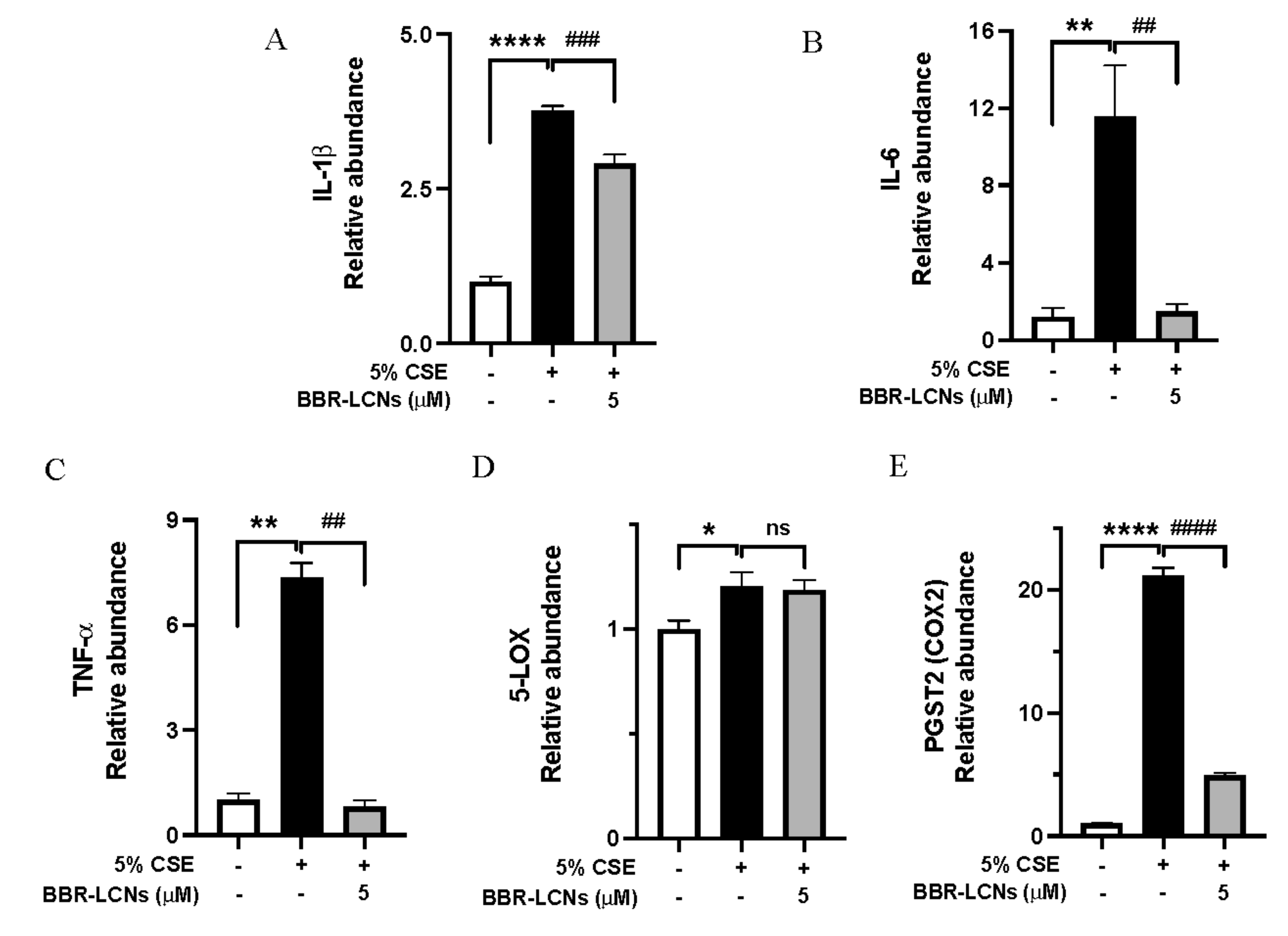
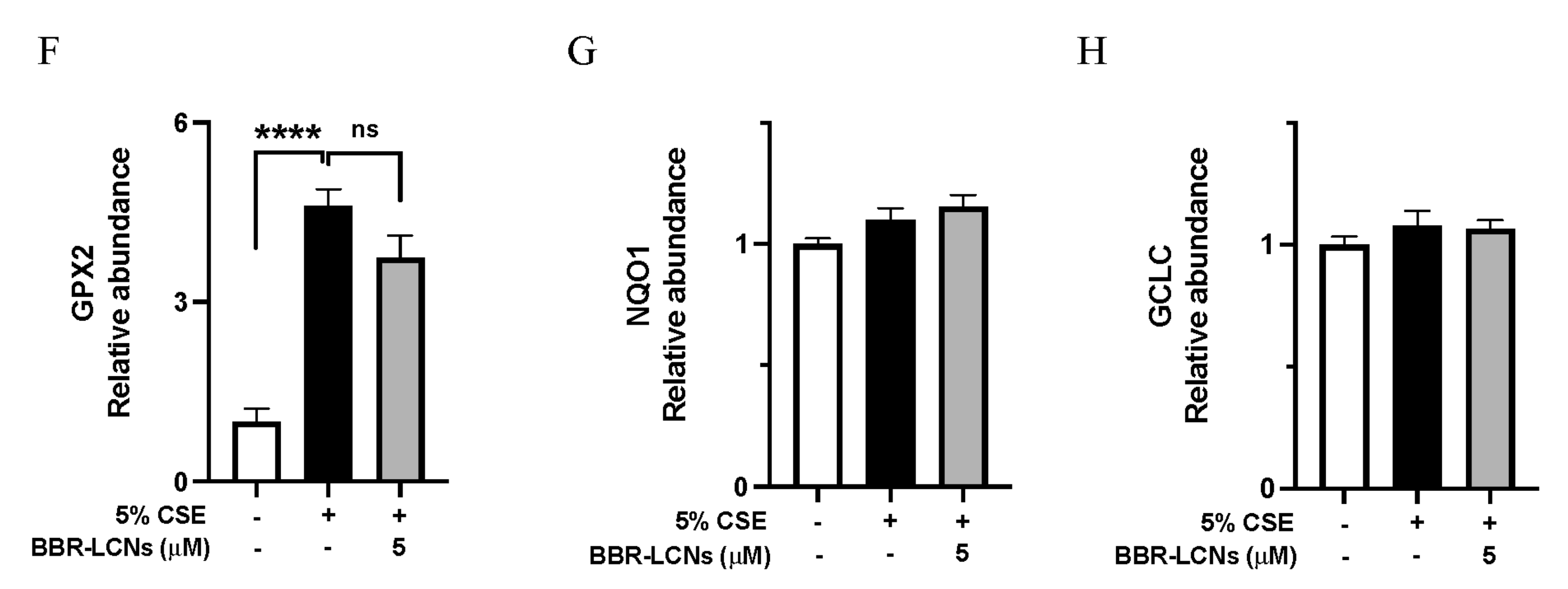
3.5. Inhibition of Inflammation and Oxidative Stress-Related Gene Expression in 16HBE Cells by BBR-LCNs
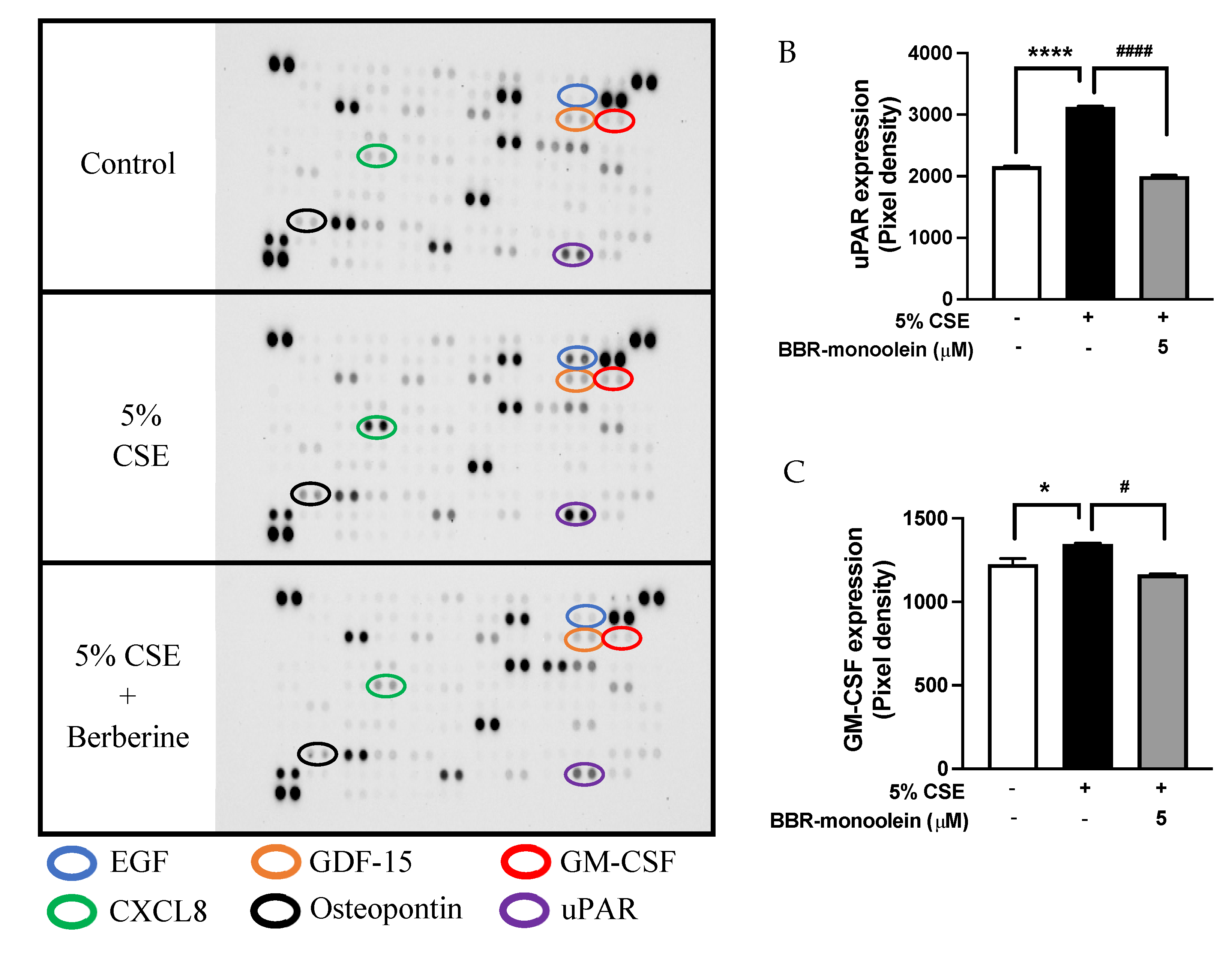
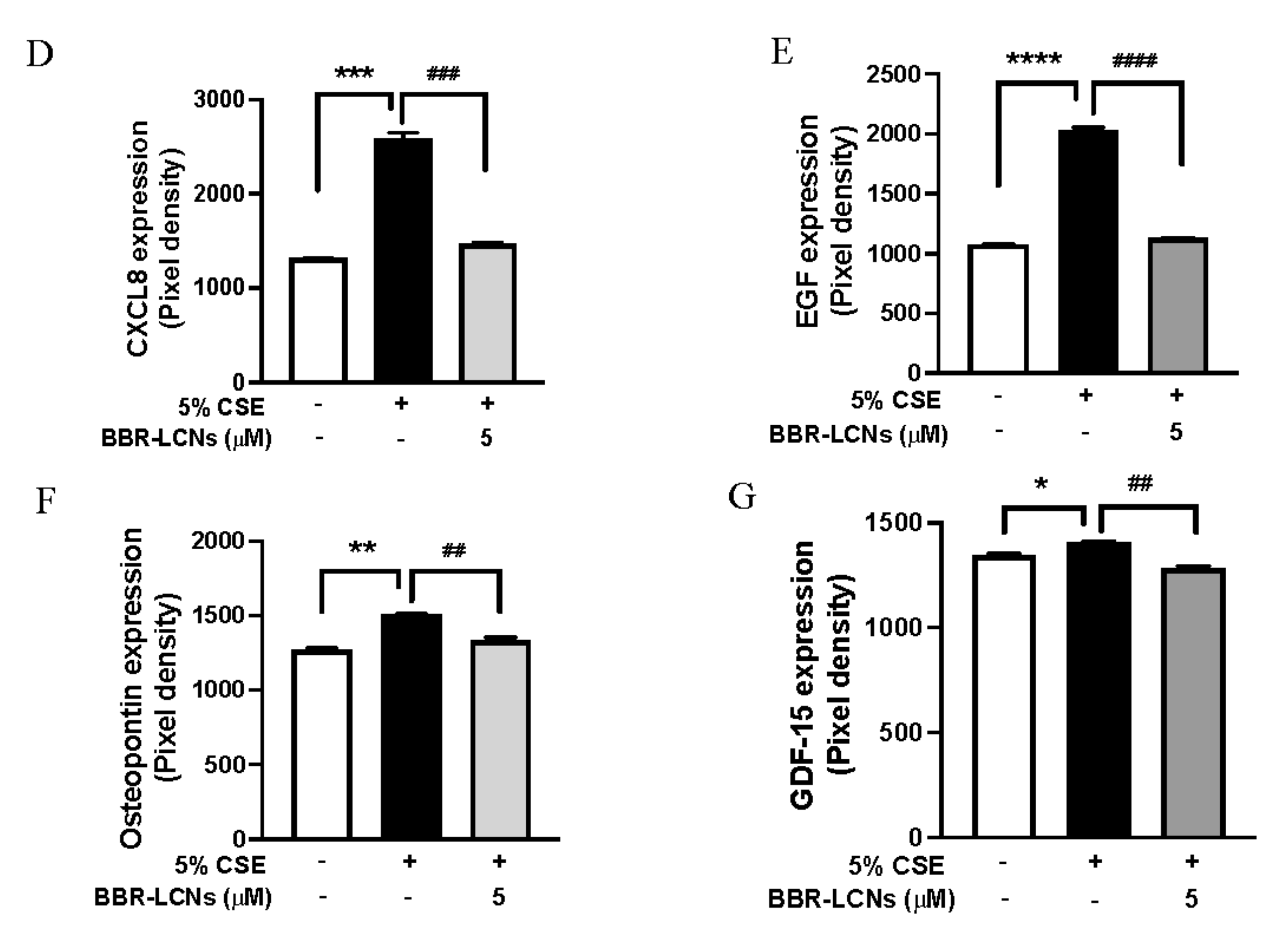
3.6. Inhibition of 5% CSE Induced Cytokines Protein Expression in 16HBE Cells by BBR-LCNs
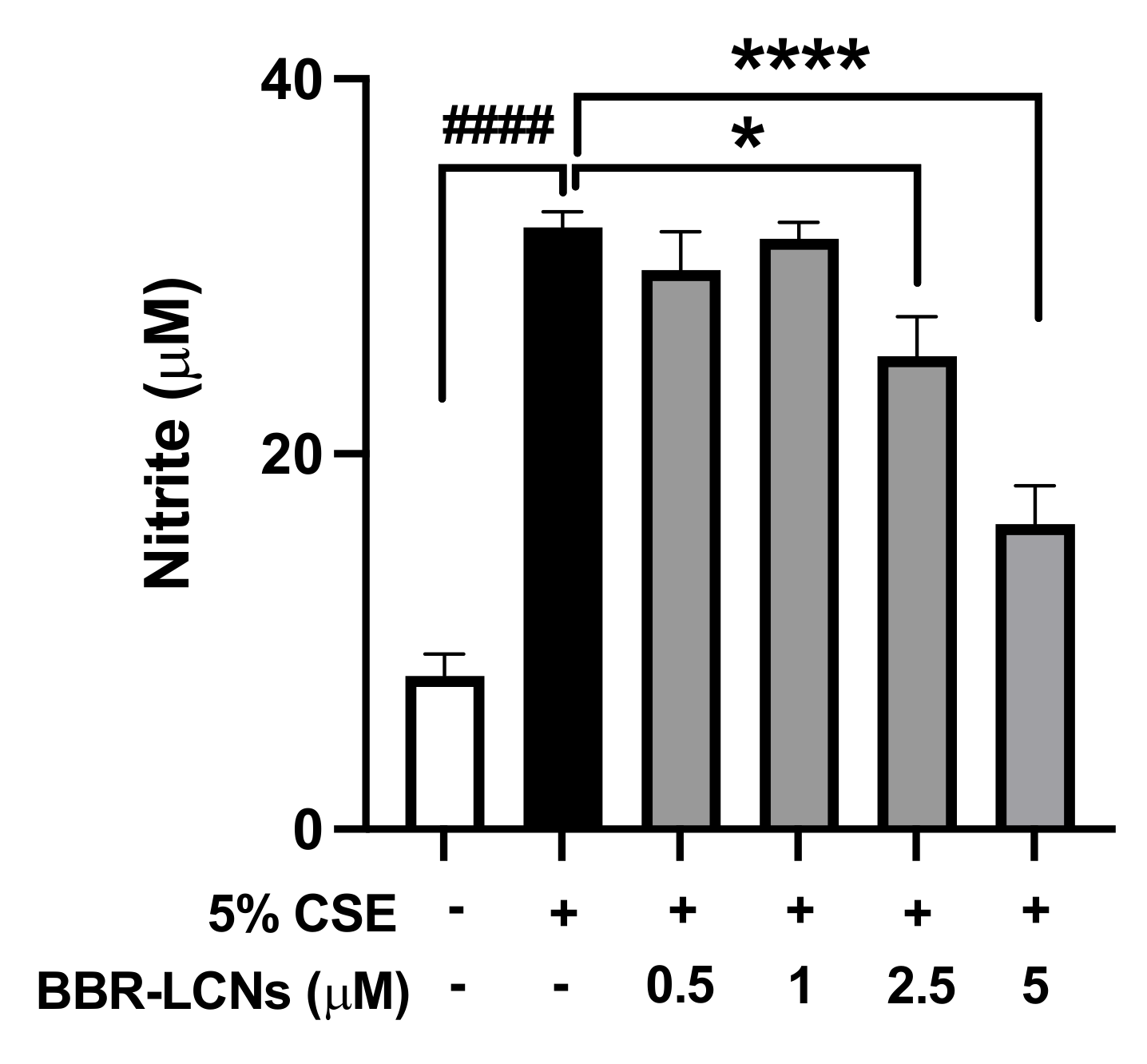
3.7. Inhibition of NO Production in RAW264.7 Cells by BBR-LCNs
3.8. Inhibition of CSE-Induced ROS Generation in RAW264.7 Cells by BBR-LCNs
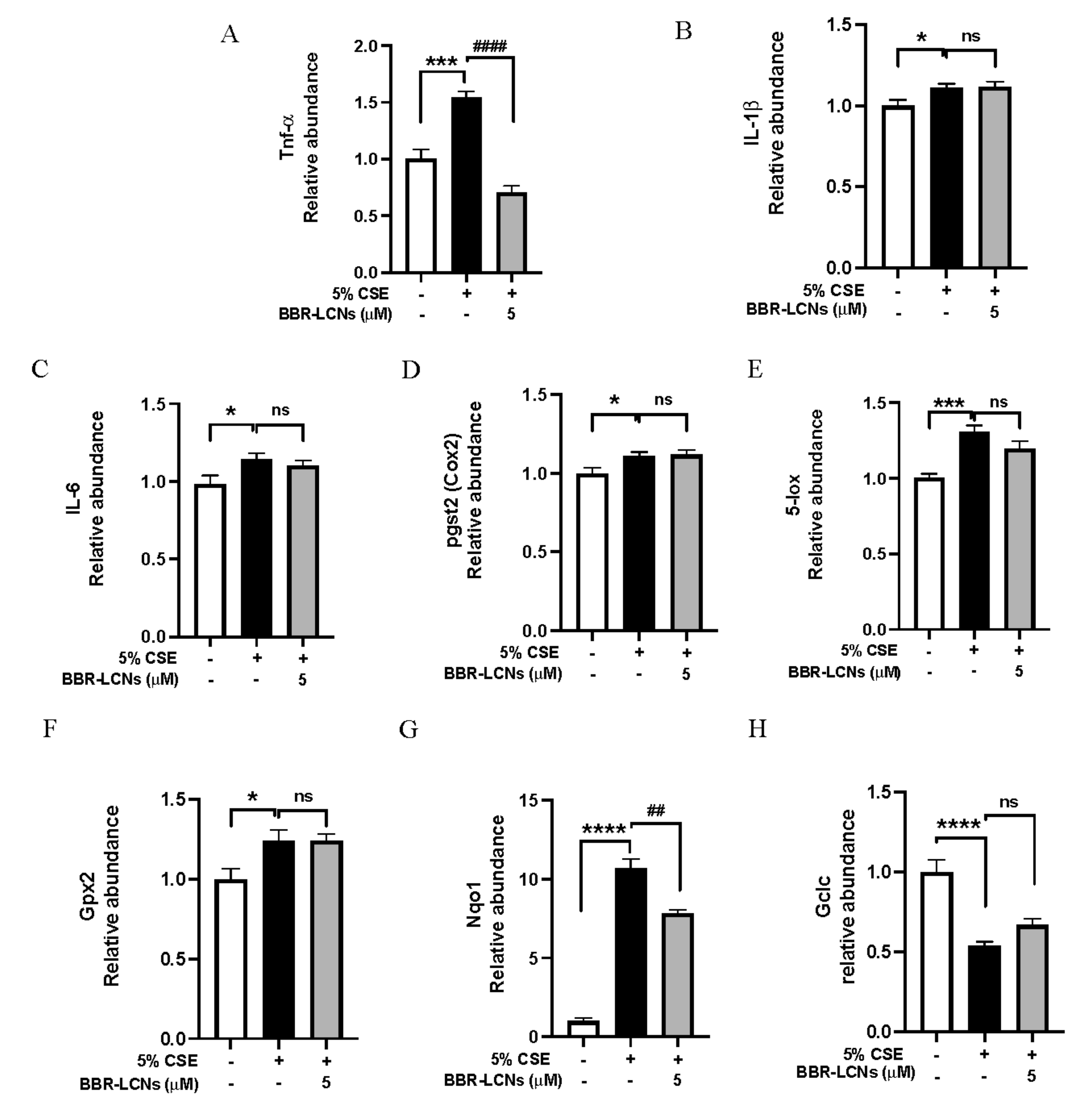
3.9. Inhibition of Oxidative Stress and Inflammation-Related Gene Expression in RAW264.7 Cells by BBR-LCNs
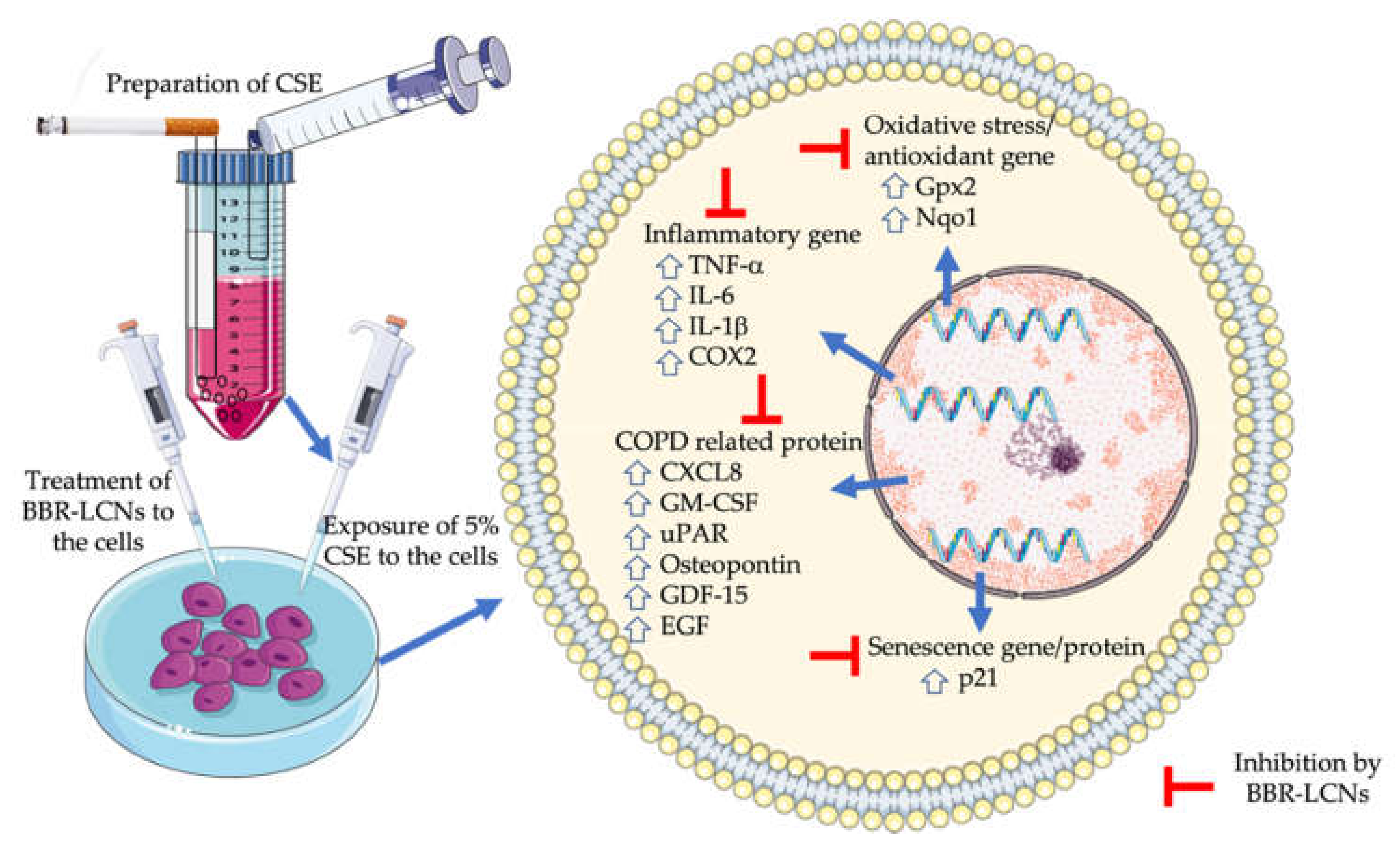
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mehta, M.; Satija, S.; Paudel, K.R.; Malyla, V.; Kannaujiya, V.K.; Chellappan, D.K.; Bebawy, M.; Hansbro, P.M.; Wich, P.R.; Dua, K. Targeting respiratory diseases using miRNA inhibitor based nanotherapeutics: Current status and future perspectives. Nanomedicine 2021, 31, 102303. [Google Scholar] [CrossRef] [PubMed]
- Paudel, K.R.; Dharwal, V.; Patel, V.K.; Galvao, I.; Wadhwa, R.; Malyla, V.; Shen, S.S.; Budden, K.F.; Hansbro, N.G.; Vaughan, A.; et al. Role of Lung Microbiome in Innate Immune Response Associated With Chronic Lung Diseases. Front. Med. (Lausanne) 2020, 7, 554. [Google Scholar] [CrossRef] [PubMed]
- Paudel, K.R.; Jha, S.K.; Allam, V.S.R.R.; Prasher, P.; Gupta, P.K.; Bhattacharjee, R.; Jha, N.K.; Vishwas, S.; Singh, S.K.; Shrestha, J.J.P. Recent Advances in Chronotherapy Targeting Respiratory Diseases. Pharmaceutics 2021, 13, 2008. [Google Scholar] [CrossRef] [PubMed]
- Malyla, V.; Paudel, K.R.; Shukla, S.D.; Donovan, C.; Wadhwa, R.; Pickles, S.; Chimankar, V.; Sahu, P.; Bielefeldt-Ohmann, H.; Bebawy, M.; et al. Recent advances in experimental animal models of lung cancer. Future Med. Chem. 2020, 12, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Guertin, K.A.; Grant, R.K.; Arnold, K.B.; Burwell, L.; Hartline, J.; Goodman, P.J.; Minasian, L.M.; Lippman, S.M.; Klein, E.; Cassano, P.A. Effect of long-term vitamin E and selenium supplementation on urine F2-isoprostanes, a biomarker of oxidative stress. Free Radic. Biol. Med. 2016, 95, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Nucera, F.; Mumby, S.; Paudel, K.R.; Dharwal, V.; Casolaro, V.; Hansbro, P.M.; Adcock, I.M.; Caramori, G. Role of oxidative stress in the pathogenesis of COPD. Minerva Med. 2022. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiao, W.; Zhang, Y.; Xing, Y. Urokinase-type plasminogen activator system and human cationic antimicrobial protein 18 in serum and induced sputum of patients with chronic obstructive pulmonary disease. Respirology 2010, 15, 939–946. [Google Scholar] [CrossRef]
- De Boer, W.I.; Hau, C.M.; van Schadewijk, A.; Stolk, J.; van Krieken, J.H.; Hiemstra, P.S. Expression of epidermal growth factors and their receptors in the bronchial epithelium of subjects with chronic obstructive pulmonary disease. Am. J. Clin. Pathol. 2006, 125, 184–192. [Google Scholar] [CrossRef]
- Mueller, T.; Leitner, I.; Egger, M.; Haltmayer, M.; Dieplinger, B. Association of the biomarkers soluble ST2, galectin-3 and growth-differentiation factor-15 with heart failure and other non-cardiac diseases. Clin. Chim. Acta 2015, 445, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.N.; Mori, M.; Mertens, T.C.; Siddhuraj, P.; Erjefält, J.S.; Önnerfjord, P.; Hiemstra, P.S.; Egesten, A.J.S.R. Osteopontin expression in small airway epithelium in COPD is dependent on differentiation and confined to subsets of cells. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Wu, J.; Li, A.; Bi, W.; Liu, T.; Cao, L.; Liu, Y.; Dong, L. The inhibitory mechanism of Cordyceps sinensis on cigarette smoke extract-induced senescence in human bronchial epithelial cells. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 1721–1731. [Google Scholar] [CrossRef] [Green Version]
- Culpitt, S.V.; Rogers, D.F.; Fenwick, P.S.; Shah, P.; De Matos, C.; Russell, R.E.; Barnes, P.J.; Donnelly, L.E. Inhibition by red wine extract, resveratrol, of cytokine release by alveolar macrophages in COPD. Thorax 2003, 58, 942–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Dong, Z.; Li, N.; Feng, X.; Liu, Y.; Li, A.; Zhu, X.; Li, C.; Zhao, Z. Nucleosides isolated from Ophiocordyceps sinensis inhibit cigarette smoke extract-induced inflammation via the SIRT1-nuclear factor-kappaB/p65 pathway in RAW264.7 macrophages and in COPD mice. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 2821–2832. [Google Scholar] [CrossRef] [Green Version]
- Mortaz, E.; Henricks, P.A.; Kraneveld, A.D.; Givi, M.E.; Garssen, J.; Folkerts, G. Cigarette smoke induces the release of CXCL-8 from human bronchial epithelial cells via TLRs and induction of the inflammasome. Biochim. Biophys. Acta 2011, 1812, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Raju Allam, V.S.R.; Paudel, K.R.; Singh, S.K.; Gulati, M.; Dhanasekaran, M.; Gupta, P.K.; Jha, N.K.; Devkota, H.P.; Gupta, G.; et al. Nutraceuticals: Unlocking newer paradigms in the mitigation of inflammatory lung diseases. Crit. Rev. Food Sci. Nutr. 2021, 10, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.M.; Paudel, K.R.; Kim, D.W. Eriobotrya japonica leaf extract attenuates airway inflammation in ovalbumin-induced mice model of asthma. J. Ethnopharmacol. 2020, 253, 112082. [Google Scholar] [CrossRef]
- Paudel, K.R.; Panth, N. Phytochemical Profile and Biological Activity of Nelumbo nucifera. Evid. Based Complement. Altern. Med. 2015, 2015, 789124. [Google Scholar] [CrossRef] [Green Version]
- Devkota, H.P.; Gaire, B.P.; Hori, K.; Subedi, L.; Adhikari-Devkota, A.; Belwal, T.; Paudel, K.R.; Jha, N.K.; Singh, S.K.; Chellappan, D.K.; et al. The science of matcha: Bioactive compounds, analytical techniques and biological properties. Trends Food Sci. Technol. 2021, 118, 735–743. [Google Scholar] [CrossRef]
- Panth, N.; Manandhar, B.; Paudel, K.R. Anticancer Activity of Punica granatum (Pomegranate): A Review. Phytother. Res. 2017, 31, 568–578. [Google Scholar] [CrossRef]
- Paudel, K.R.; Wadhwa, R.; Tew, X.N.; Lau, N.J.X.; Madheswaran, T.; Panneerselvam, J.; Zeeshan, F.; Kumar, P.; Gupta, G.; Anand, K.; et al. Rutin loaded liquid crystalline nanoparticles inhibit non-small cell lung cancer proliferation and migration in vitro. Life Sci. 2021, 276, 119436. [Google Scholar] [CrossRef]
- Mehta, M.; Paudel, K.R.; Shukla, S.D.; Shastri, M.D.; Satija, S.; Singh, S.K.; Gulati, M.; Dureja, H.; Zacconi, F.C.; Hansbro, P.M.; et al. Rutin-loaded liquid crystalline nanoparticles attenuate oxidative stress in bronchial epithelial cells: A PCR validation. Future Med. Chem. 2021, 13, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Solanki, N.; Mehta, M.; Chellappan, D.K.; Gupta, G.; Hansbro, N.G.; Tambuwala, M.M.; Aa Aljabali, A.; Paudel, K.R.; Liu, G.; Satija, S.; et al. Antiproliferative effects of boswellic acid-loaded chitosan nanoparticles on human lung cancer cell line A549. Future Med. Chem. 2020, 12, 2019–2034. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, R.; Paudel, K.R.; Chin, L.H.; Hon, C.M.; Madheswaran, T.; Gupta, G.; Panneerselvam, J.; Lakshmi, T.; Singh, S.K.; Gulati, M.; et al. Anti-inflammatory and anticancer activities of Naringenin-loaded liquid crystalline nanoparticles in vitro. J. Food Biochem. 2021, 45, e13572. [Google Scholar] [CrossRef] [PubMed]
- Paudel, K.R.; Wadhwa, R.; Mehta, M.; Chellappan, D.K.; Hansbro, P.M.; Dua, K. Rutin loaded liquid crystalline nanoparticles inhibit lipopolysaccharide induced oxidative stress and apoptosis in bronchial epithelial cells in vitro. Toxicol. In Vitro 2020, 68, 104961. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Kaushik, N. Phytochemical and pharmacological studies in genus Berberis. Phytochem. Rev. 2012, 11, 523–542. [Google Scholar] [CrossRef]
- Ma, J.; Chan, C.C.; Huang, W.C.; Kuo, M.L. Berberine Inhibits Pro-inflammatory Cytokine-induced IL-6 and CCL11 Production via Modulation of STAT6 Pathway in Human Bronchial Epithelial Cells. Int. J. Med. Sci. 2020, 17, 1464–1473. [Google Scholar] [CrossRef]
- Gupta, S.; Khan, A.; Vishwas, S.; Gulati, M.; Gurjeet Singh, T.; Dua, K.; Kumar Singh, S.; Najda, A.; Sayed, A.A.; Almeer, R.; et al. Demethyleneberberine: A possible treatment for Huntington’s disease. Med. Hypotheses 2021, 153, 110639. [Google Scholar] [CrossRef]
- Spinozzi, S.; Colliva, C.; Camborata, C.; Roberti, M.; Ianni, C.; Neri, F.; Calvarese, C.; Lisotti, A.; Mazzella, G.; Roda, A. Berberine and its metabolites: Relationship between physicochemical properties and plasma levels after administration to human subjects. J. Nat. Prod. 2014, 77, 766–772. [Google Scholar] [CrossRef]
- Liu, C.-S.; Zheng, Y.-R.; Zhang, Y.-F.; Long, X.-Y. Research progress on berberine with a special focus on its oral bioavailability. Fitoterapia 2016, 109, 274–282. [Google Scholar] [CrossRef]
- Ng, P.Q.; Ling, L.S.C.; Chellian, J.; Madheswaran, T.; Panneerselvam, J.; Kunnath, A.P.; Gupta, G.; Satija, S.; Mehta, M.; Hansbro, P.M.; et al. Applications of Nanocarriers as Drug Delivery Vehicles for Active Phytoconstituents. Curr. Pharm. Des. 2020, 26, 4580–4590. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Advances in nanomedical applications: Diagnostic, therapeutic, immunization, and vaccine production. Environ. Sci. Pollut. Res. Int. 2020, 27, 19200–19213. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Mehta, M.; Paudel, K.R.; Madheswaran, T.; Panneerselvam, J.; Gupta, G.; Su, Q.P.; Hansbro, P.M.; MacLoughlin, R.; Dua, K.; et al. Versatility of liquid crystalline nanoparticles in inflammatory lung diseases. Nanomedicine 2021, 16, 1545–1548. [Google Scholar] [CrossRef] [PubMed]
- Paudel, K.R.; Mehta, M.; Yin, G.H.S.; Yen, L.L.; Malyla, V.; Patel, V.K.; Panneerselvam, J.; Madheswaran, T.; MacLoughlin, R.; Jha, N.K.; et al. Berberine-loaded liquid crystalline nanoparticles inhibit non-small cell lung cancer proliferation and migration in vitro. Environ. Sci. Pollut. Res. Int. 2022, 1–18. [Google Scholar] [CrossRef]
- Mehta, M.; Malyla, V.; Paudel, K.R.; Chellappan, D.K.; Hansbro, P.M.; Oliver, B.G.; Dua, K. Berberine loaded liquid crystalline nanostructure inhibits cancer progression in adenocarcinomic human alveolar basal epithelial cells in vitro. J. Food Biochem. 2021, 45, e13954. [Google Scholar] [CrossRef] [PubMed]
- Leus, N.G.; van den Bosch, T.; van der Wouden, P.E.; Krist, K.; Ourailidou, M.E.; Eleftheriadis, N.; Kistemaker, L.E.; Bos, S.; Gjaltema, R.A.; Mekonnen, S.A.; et al. HDAC1-3 inhibitor MS-275 enhances IL10 expression in RAW264.7 macrophages and reduces cigarette smoke-induced airway inflammation in mice. Sci. Rep. 2017, 7, 45047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijink, I.H.; Brandenburg, S.M.; Postma, D.S.; van Oosterhout, A.J. Cigarette smoke impairs airway epithelial barrier function and cell-cell contact recovery. Eur. Respir. J. 2012, 39, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-H.; Paudel, K.R.; Kim, D.-W. Terminalia chebula fructus inhibits migration and proliferation of vascular smooth muscle cells and production of inflammatory mediators in RAW 264.7. Evid.-Based Complement. Altern. Med. 2015, 2015, 1–10. [Google Scholar]
- Paudel, K.R.; Karki, R.; Kim, D.W. Cepharanthine inhibits in vitro VSMC proliferation and migration and vascular inflammatory responses mediated by RAW264.7. Toxicol. In Vitro 2016, 34, 16–25. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, P.; Hanaoka, M.; Droma, Y.; Kubo, K. Enhanced levels of prostaglandin E2 and matrix metalloproteinase-2 correlate with the severity of airflow limitation in stable COPD. Respirology 2008, 13, 1014–1021. [Google Scholar] [CrossRef]
- Kennedy-Feitosa, E.; Pinto, R.F.; Pires, K.M.; Monteiro, A.P.; Machado, M.N.; Santos, J.C.; Ribeiro, M.L.; Zin, W.A.; Canetti, C.A.; Romana-Souza, B.; et al. The influence of 5-lipoxygenase on cigarette smoke-induced emphysema in mice. Biochim. Biophys. Acta 2014, 1840, 199–208. [Google Scholar] [CrossRef]
- Gogtay, N.J.; Bhatt, H.A.; Dalvi, S.S.; Kshirsagar, N.A. The use and safety of non-allopathic Indian medicines. Drug Saf. 2002, 25, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Tamimi, A.; Serdarevic, D.; Hanania, N.A. The effects of cigarette smoke on airway inflammation in asthma and COPD: Therapeutic implications. Respir. Med. 2012, 106, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preethi, S.; Arthiga, K.; Patil, A.B.; Spandana, A.; Jain, V. Review on NAD(P)H dehydrogenase quinone 1 (NQO1) pathway. Mol. Biol. Rep. 2022, 1–18. [Google Scholar] [CrossRef]
- Fredenburgh, L.E.; Perrella, M.A.; Mitsialis, S.A. The role of heme oxygenase-1 in pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2007, 36, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Kondo, N.; Cano, M.; Ebrahimi, K.; Handa, J.J.I.O.; Science, V. Nrf2 Signaling is Activated After Cigarette Smoke Extract Exposure in Rpe Cells in vitro and in vivo. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4104. [Google Scholar]
- Bazzini, C.; Rossetti, V.; Civello, D.A.; Sassone, F.; Vezzoli, V.; Persani, L.; Tiberio, L.; Lanata, L.; Bagnasco, M.; Paulmichl, M.; et al. Short- and long- term effects of cigarette smoke exposure on glutathione homeostasis in human bronchial epithelial cells. Cell Physiol. Biochem. 2013, 32, 129–145. [Google Scholar] [CrossRef] [Green Version]
- Obeidat, M.; Dvorkin-Gheva, A.; Li, X.; Bosse, Y.; Brandsma, C.A.; Nickle, D.C.; Hansbro, P.M.; Faner, R.; Agusti, A.; Pare, P.D.; et al. The Overlap of Lung Tissue Transcriptome of Smoke Exposed Mice with Human Smoking and COPD. Sci. Rep. 2018, 8, 11881. [Google Scholar] [CrossRef] [Green Version]
- Pickett, G.; Seagrave, J.; Boggs, S.; Polzin, G.; Richter, P.; Tesfaigzi, Y. Effects of 10 cigarette smoke condensates on primary human airway epithelial cells by comparative gene and cytokine expression studies. Toxicol. Sci. 2010, 114, 79–89. [Google Scholar] [CrossRef]
- Rumzhum, N.N.; Ammit, A.J. Cyclooxygenase 2: Its regulation, role and impact in airway inflammation. Clin. Exp. Allergy 2016, 46, 397–410. [Google Scholar] [CrossRef]
- Kilfeather, S. 5-lipoxygenase inhibitors for the treatment of COPD. Chest 2002, 121, 197S–200S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Kim, N.-H.; Kim, K.-S. Berberine Suppresses MUC5AC Gene in Airway Epithelial Cells. Otolaryngol.–Head Neck Surg. 2010, 143, P269–P270. [Google Scholar] [CrossRef]
- Gong, M.; Duan, H.; Wu, F.; Ren, Y.; Gong, J.; Xu, L.; Lu, F.; Wang, D. Berberine Alleviates Insulin Resistance and Inflammation via Inhibiting the LTB4-BLT1 Axis. Front. Pharmacol. 2021, 12, 722360. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.A.; Cheon, H.G. Activating transcription factor-3 induction is involved in the anti-inflammatory action of berberine in RAW264.7 murine macrophages. Korean J. Physiol. Pharmacol. 2016, 20, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckett, E.L.; Stevens, R.L.; Jarnicki, A.G.; Kim, R.Y.; Hanish, I.; Hansbro, N.G.; Deane, A.; Keely, S.; Horvat, J.C.; Yang, M.; et al. A new short-term mouse model of chronic obstructive pulmonary disease identifies a role for mast cell tryptase in pathogenesis. J. Allergy Clin. Immunol. 2013, 131, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Singanayagam, A.; Glanville, N.; Walton, R.P.; Aniscenko, J.; Pearson, R.M.; Pinkerton, J.W.; Horvat, J.C.; Hansbro, P.M.; Bartlett, N.W.; Johnston, S.L. A short-term mouse model that reproduces the immunopathological features of rhinovirus-induced exacerbation of COPD. Clin. Sci. 2015, 129, 245–258. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Gene Name | Forward Sequence | Reverse Sequence | Accession Number (RefSeqID) |
|---|---|---|---|---|
| 16HBE | SIRT1 | AAGGAAAACTACTTCGCAAC | GGAACCATGACACTGAATTATC | NM_001142498 |
| CDKN1A(p21) | CAGCATGACAGATTTCTACC | CAGGGTATGTACATGAGGAG | NM_000389 | |
| GPX2 | AATTTGGACATCAGCTGC | GGCTGCTCTTCAAGATTTAG | NM_000852 | |
| IL-1β | GCCTCAAGGAAAAGAATCTG | GGATCTACACTCTCCAGG | NM_000576 | |
| IL-6 | GCAGAAAAAGGCAAAGAATC | CTACATTTGCCGAAGAGC | NM_000600 | |
| TNF-α | AGGCAGTCAGATCATCTTC | TTATCTCTCAGCTCCACG | NM_000594 | |
| NQO-1 | AGTATCCACAATAGCTGACG | TTTGTGGGTCTGTAGAAATG | NM_000903 | |
| GCLC | TTATTAGAGACCCACTGACAC | TTCTCAAAATGGTCAGACTC | NM_001197115 | |
| PTGS2 (COX-2) | AAGCAGGCTAATACTGATAGG | TGTTGAAAAGTAGTTCTGGG | NM_000963 | |
| 5-LOX | AAATGCCACAAGGATTTACC | ATCGCTTTGGAGTAATTCAG | NM_000698 | |
| GAPDH | TCGGAGTCAACGGATTTG | CAACAATATCCACTTTACCAGAG | NM_002046 | |
| RAW264.7 | Gclc | CGACCAATGGAGGTGCAGTTA | AACCTTGGACAGCGGAATGA | NM_010295.2 |
| Nqo1 | GTAGCGGCTCCATGTACTCTC | AGGATGCCACTCTGAATCGG | NM_008706.5 | |
| Tnf-α | TCTGTCTACTGAACTTCGGGGTGA | TTGTCTTTGAGATCCATGCCGTT | NM_013693.3 | |
| IL-6 | AGAAAACAATCTGAAACTTCCAGAGAT | GAAGACCAGAGGAAATTTTCAATAGG | NM_031168 | |
| Gpx2 | ACC AGTTCGGACATCAGGAG | CCC AGGTCGGACATACTTGA | NM_030677 | |
| IL-1β | TGGGATCCTCTCCAGCCAAGC | AGCCCTTCATCTTTTGGGGTCCG | NM_008361 | |
| Ptgs2 (Cox-2) | ACTCATAGGAGAGACTATCAAG | GAGTGTGTTGAATTCAGAGG | NM_011198 | |
| Alox5 (5-Lox) | CAGGAAGGGAACATTTTCATC | AGGAAGATTGGGTTACTCTC | NM_009662 | |
| Hprt | AGGCCAGACTTTGTTGGATTTGAA | CAACTTGCGCTCATCTTAGGCTTT | NM_013556.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paudel, K.R.; Panth, N.; Manandhar, B.; Singh, S.K.; Gupta, G.; Wich, P.R.; Nammi, S.; MacLoughlin, R.; Adams, J.; Warkiani, M.E.; et al. Attenuation of Cigarette-Smoke-Induced Oxidative Stress, Senescence, and Inflammation by Berberine-Loaded Liquid Crystalline Nanoparticles: In Vitro Study in 16HBE and RAW264.7 Cells. Antioxidants 2022, 11, 873. https://doi.org/10.3390/antiox11050873
Paudel KR, Panth N, Manandhar B, Singh SK, Gupta G, Wich PR, Nammi S, MacLoughlin R, Adams J, Warkiani ME, et al. Attenuation of Cigarette-Smoke-Induced Oxidative Stress, Senescence, and Inflammation by Berberine-Loaded Liquid Crystalline Nanoparticles: In Vitro Study in 16HBE and RAW264.7 Cells. Antioxidants. 2022; 11(5):873. https://doi.org/10.3390/antiox11050873
Chicago/Turabian StylePaudel, Keshav Raj, Nisha Panth, Bikash Manandhar, Sachin Kumar Singh, Gaurav Gupta, Peter R. Wich, Srinivas Nammi, Ronan MacLoughlin, Jon Adams, Majid Ebrahimi Warkiani, and et al. 2022. "Attenuation of Cigarette-Smoke-Induced Oxidative Stress, Senescence, and Inflammation by Berberine-Loaded Liquid Crystalline Nanoparticles: In Vitro Study in 16HBE and RAW264.7 Cells" Antioxidants 11, no. 5: 873. https://doi.org/10.3390/antiox11050873